معرفی سایت های مفید
آموزش ژنتیک
مجله ی وراثت
:http://jhered.oxfordjournals.org/
مجله ی ژنتیک ملکولی انسان از سری مجلات دانشگاه آکسفورد
:http://hmg.oxfordjournals.org/
وبسایت ژن و تکامل ارائه کننده ی بایگانی کاملی از مقالات در این زمینه از سال 1987
بخش ژنتیک مجله ی نیچر
:http://nature.com/ng/index.html
مجله ی آمریکایی ژنتیک انسانی زیر نظر دانشگاه شیکاگو، ارائه کننده ی آخرین اخبار
http://journals.uchicago.edu/AJHG/home.html
نسخه ی رسمی و حرفه ای مجله(ژورنال) اروپایی ژنتیک انسانی که در واقع بخشی
http://nature.com/ejhg/index.html
مرکز آموزش دانش ژنتیک و اخبار و تازه های این رشته به زبان انگلیسی:
انیمیشنهای زیبای زیست شناسی
ژن جدید اکسیر جوانی ابدی کشف شد
دانشمندان انگلیسی اعلام کردند که با آزمایش بر روی کرمها موفق به کشف ژنی شده اند که پیوند محکمی با طول عمر دارد و می تواند به عنوان یکی از ژنهای اکسیر جوانی ابدی معرفی شود.
بدانشمندان دانشگاه بیرمنگام که بر روی ژنتیک پیری مطالعه می کنند دریافتند که ژنی به نام DAF-16 که علاوه بر کرمها در بسیاری از حیوانات و انسان نیز وجود دارد می تواند سناریوی جدیدی در روش مبارزه با پیری و مسائل ایمنی و دفاع انسان بگشاید.
جمعیتهای جهانی از یک سو به دلیل بهبود خدمات بهداشتی و از سوی دیگر به دلیل کاهش تولد با یک ریتم بسیار سریع به سوی پیر شدن پیش می روند. نتایج تحقیقاتی که سال گذشته محققان دانمارکی انجام دادند نشان می دهد که نیمی از کودکانی که در عصر حاضر متولد می شوند می توانند تا 100 سالگی عمر کنند.
این محققان در این خصوص اظهار داشتند: "ما می خواستیم کشف کنیم که پیری طبیعی چقدر به ژنها وابسته است و اثرات این ژنها تا چه حد بر روی سایر ویژگیها مثل ایمنی اثر می گذارند. ما کشف کردیم که مسائلی از قبیل مقاومت و پیری گامهایی را به موازات هم بر می دارند."
دانشمندان انگلیسی با مقایسه طول عمر، مقاومت در برابر استرس و سیستم ایمنی در چهار گونه مختلف کرمها نشان دادند که در بین این گونه ها تفاوتهای قابل توجهی در رفتار ژن DAF-16 وجود دارد که این تفاوتها منجر به ایجاد تفاوتهایی در طول عمر، مقاومت در برابر سختیها و سیستم ایمنی این گونه ها می شود به طوری که در سطح حداکثر فعالیت DAF-16، مدت زمان زندگی و به موازات آن مقاومت و دفاع ایمنی در مقابل برخی ازعفونتها افزایش می یابد.
DAF-16 در بسیاری از سلولهای بدن فعال است و شباهت بسیاری به خانواده ای از ژنهای انسان به نام FOXO دارد.
براساس گزارش رویترز، این پژوهشگران اظهار داشتند: "تفاوتهای ناچیزی در شدت فعالیت DAF-16 در گونه های مختلف وجود دارد که به نظر می رسد این تفاوتهای جزئی نقش مهمی در پیری و سلامت ایفا کنند که بسیار جالب است و می تواند توضیح دهد که با دستکاری این ژن می توان بر روی طول عمر تحولات جدی ایجاد کرد."
انتقال ويروس ايدز از طريق اسپرم

آنا سبالوس محقق برجسته و معروف دانشگاه بوئينوس آيرس در آرژانتين پس از انجام آزمايشات جديد خود ادعا كرده است كه اسپرم هم ميتواند ويروس اچ.آي.وي را به ماكروفاژها، سلولهايT شكل و سلولهاي دندريت منتقل كند.
دكتر سبالوس و دستياران وي نشان دادند كه ويروس اچ.آي.وي به سطح اسپرم ميچسبد و اين اسپرمهاي حامل ويروس، آن را بهدندريتها و ساير سلولها و بافتهاي مورد هدف ويروس ايدز منتقل ميكند.
اسپرم بيانكننده و حامل مولكولهايي است كه در تعامل براي جاسازي ويروس ايدز نقش دارند، از جمله سولفات هپارين و گيرندههاي مانوس كه در انتشار ويروس ايدز در بدن نقش ايفا ميكنند. اين محققان نشان دادند كه ويروس اچ.آي.وي از راه سولفات هپارين به اسپرم متصل ميشود، اما گيرندههاي مانوس در عملكرد اسپرمي تاثيري ندارند. آزمايشات نشان داد: وقتي ويروس ايدز به اسپرم متصل شد در محيط كشت بافت از طريق اسپرم به سلولهاي دندريت ميرسد، سپس سلولهاي دندريت در تعامل با ويروس در بدن سبب بروز التهاب ميشود.
محققان معتقدند كه پروسه تضعيف سيستم ايمني بدن توام با بروز اين التهاب، انتشار ويروس ايدز را در بدن تشديد ميكند.
كشف روشي براي خشك كردن موقت سلولها با قابليت احياي مجدد
دانشمندان چك با همكاري دستياران ايتاليايي خود موفق به كشف روش جديدي براي ذخيره و نگهداري سلولها شدند تا بتوان در مواقع ضرورت آنها را احيا كرد.
به گزارش سرويس «علمي» خبرگزاري دانشجويان ايران (ايسنا)، پژوهشگران پراگ و ساردينيا اولين محققان در جهان هستند كه روش خشك كردن موقتي سلولها را براي زنده كردن دوباره آنها در صورت لزوم كشف كردهاند.
براساس گزارش كريستن ساينس مانيتور، پژوهشگران از اين روش در شبيه سازي جنينهاي گوسفند استفاده كردند.
با اين كشف به زودي پژوهشگران ميتوانند يك بانك اطلاعاتي از سلولهاي خشك شده متعلق به گونه هاي جانوري رو به انقراض تاسيس كنند. به اين ترتيب ميتوان از اين ذخيره براي شبيه سازي اين جانوران در آينده استفاده كرد.
اين مطالعات در موسسه تحقيقات توليدات حيواني فرهنگستان علوم چك انجام شده است.
براساس اين گزارش، تاكنون اسپرم تنها سلول داراي هسته بوده كه دانشمندان ذخيره آن را در وضعيت دهتيراته (بدون آب) كنترل كردهاند.
پژوهشگران در آينده ميتوانند از اين روش براي كمك به احياي اعضاي بدن كه مورد حمله سرطانها قرار گرفتهاند به استفاده نمايند.
قند ضد یخ درون سلولی
استفاده از موجهاى تراهرتز براى رديابى واكنشهاى درون سلولى
ایتفاده ازطول موجهاى مختلف در ساخت لوازم خانگى و زندگى روزمره عادى شده است. غذا را مىتوان به سرعت در ميكرووله گرم كرد و يا تلويزيون را از راه دور كنترل كرد. اما دقيقأ بين اين دو نوع مختلف طول موج، فركانسى قرار دارد كه تا كنون از آن استفاده تكنيكى نشده است: موجهاى تراهرتز Terahertz-Wellen. هر روز دامنه تحقيقات در زمينه نحوه استفاده از امواج تراهرتز گستردهتر مىشود.بزودى شاهد لامپها و دوربينهايى خواهيم بود كه به كمك امواج تراهترز عمل خواهند كرد.
فيزيكدانان دانشگاه بوخوم در آلمان سعى مىكنند با كمك اين امواج چگونهگى و نحوه عملكرد فعاليتهاى زيستى را رديابى كنند. يكى از موضوعاتى كه از مدتى پيش در دست تحقيق است، بررسى خواص قند در فعاليتهاى حياتى است. قند نه تنها انرژىزاست بلكه نقش مهمى نيز در حفظ تعادل فعاليتهاى زيستى بر عهده دارد. مثلأ سفيده تخممرغى كه با كمى قند مخلوط شده است، ديرتر خشك مىشود و حتى ديرتر هم يخ مىبندد. به عبارت سادهتر قند مانند ضديخ بيولوژيكى عمل مىكند.
مكانيسمى كه از مدتها پيش شناخته شده است. اما چگونه؟ اين پروسه جالب كاملأ ناشناخته است.
مارتينا هاونيت (Martina Havenith) يكى از فيزيكدان دانشگاه شهر بوخوم مىگويد:
”نظرات متفاوتى در اين زمينه وجود دارد، مثلاً مىتوان گفت ملكولهاى قند مىتوانند بين ملكولهاى آب سر بخورند و جايگزين شوند و يا شايد حضور قند روى خواص آب تأثير مىگذارد و حركت ملكولهاى آنرا كند مىكند.“
براى بررسى بيشتر و اثبات اين تئوريها ابتدا بايد بتوان حركات ملكولهاى تشكيل دهنده آب را بررسى كرد. اما اين ملكولها در مدت زمانى كمتر از يك بيليونيوم ثانيه جابهجا مىشود! سرعتى برق آسا كه به سادگى قابل رديابى نيست. روشهاى كه امروزه براى رديابى حركات ملكولى استفاده مىشوند بسيار كندتر از آن هستند كه بتوانند تصويرى از نوع حركت ملكولهاى آب نشان دهند. هاونيت معتقد است مىتوان از امواج تراهرتز در تصوير بردارى از اين حركات برق آسا استفاده كرد.
ليز يا سنسورهاى امواج تراهرتز آنقدر سريع هستند كه مىتوانند اين نوع حركات ملكولى را دنبال كنند. فيزيدانان بوخومى تلاش مىكنند از اين تكنولوژى براى رديابى و نحوه عمل ملكولهاى قند استفاده كنند. بررسىهاى صورت گرفته نشان مىدهند هر ملكول قند حدود صد وده ملكول آب اطرافش را تحت تأثير قرار ميدهد. ايجاد پيوند هيدروژنى ملكولهاى آب با قند، نوعى جاذبه ايجاد مىكند. به علت اين كشش ذرههاى آب اطراف قند نمىتوانند آزادانه به هر سو حركت كنند. اما چرا اين پيوند محكم ميان قند و آب از خشك شدن و يا يخ زدن سفيده تخممرغ جلوگيرى مىكند؟ فيزيكدان آلمانى هم نمىدانند.
هاوينت مىگويد: ”همه چيز هنوز در حد تئورى است. حتى مىتواند اينگونه باشد كه چون حركت ملكولى آب كندتر مىشود، تغييرات پروتئينى هم كندتر مىشود.“
اما در هر حال اين نظريه ثابت مىكند كه حضور آب در سلولهاى زنده حالت خنثى ندارد بلكه آب بصورت فعال در تمامى روندهاى سلولى شركت مىكند و نقش تعيين كنندهاى بر عهده دارد. هاوينت:
” تعريف تصويرى اين پروسه مانند اين است ملكولهاى آب را مثل بازيكنان فوتبال در نظر بگيريم كه از بيرون به درون ضربه مىزنند و يك پروسه مشخص را راه مىاندازند حال اگر اين ضربه آرامتر باشد پروسه هم آرامتر راه مىافتد.“
دقيقأ مانند خرسى كه در زمستان به خواب زمستانى فرو مىرود چون سوخت و ساز درون سلولىاش كندتر شده است. سفيده تخممرغ هم به علت كند شدن حركت ملكولهاى آب ديرتر يخ مىزند، ديرتر هم خشك مىشود. دست تئورى فيزيكى اينگونه به نظر مىرسد.
اما شايد روزى بتوان پس از شناخت مكانيسم اين روند، ضديخ قندى توليد كرد و يا از اين روش براى ساخت داروهاى جديد از آن استفاده كرد.
شبنم نوريان/رادیو صدای المان
چرا تعداد خانم ها بيش از آقايان است؟
حتما شنيده ايد كه كروموزوم Y در حال زوال است ، ولي آيا مردان رو به نابودي اند؟ زنان بخاطر داشتن تفاوتي سيتوژنتيكي از مردان متمايزند. يك كروموزوم X اضافي بجاي Y باعث شده است آنها زن باشند.

تقريباً يك دهه از زمان اولين پيشگويي دانشمندان در رابطه با از دست رفتن كروموزوم Y در مردان مي گذرد. در واقع كروموزوم Y باعث مي شود كه مردان ، مرد باشند. اما نقشه برداري اخير از ژنوم شامپانزه نشان داده است كه از كروموزوم Y محافظت مي شود. اين امر بخاطر تك همسري انسان است.
ژن هاي انسان در 23 جفت كروموزوم گسترش يافته اند كه 22 تاي آنها كاملاً مشابه مي باشند. كروموزوم 23 همانند يك سكه ي دو رويه در عالم بيولوژي محسوب مي گردد.
دو X بمعني زن بودن و X در كنار Y بمعني مرد بودن است. جفت هاي كروموزومي غالباً قطعاتي از DNA را طي فرايند نوتركيبي ، مبادله مي كنند. نقش نوتركيبي فعال نگه داشتن ژن ها مي باشد.
تا اواخر دهه 1990 عقيده بر اين بود كه كروموزوم Y انسان بخش عمده اي از ژن هاي كروموزوم X را ندارد تا بتواند با آن تبادل قطعه انجام دهد. نتيجه اين امر باعث كاهش و غير فعال شدن تدريجي ژن هاي كروموزوم Y مي گردد تا اينكه نهايتاً محو شود. بطوريكه پيشبيني شد نسل مرد بين 5 تا 10 مليون سال اينده از كره خاكي محو خواهد شد.
چون انسان و شامپانزه از يك جد مشترك در 6 مليون سال پيش منشا گرفته اند ، ديويد پيج (David Page) از مؤسسه Whitehhead در ماساچوست بر آن شد تا روند نمو كروموزوم Y در شامپانزه و همتاي انساني آن را مطالعه نمايد. آنچه او كشف كرد دور از تصور بود:
كروموزوم Y در شامپانزه بيشتر از Y ي انساني تحليل رفته بود.
پيج و همكارانش نتيجه گرفتند كه شامپانزه هاي ماده بعلت بي بند و باري و داشتن چندين شريك جنسي ، توانسته اند ژن هاي Y را به گونه اي القا كنند كه اسپرم هاي فعالي را توليد نمايند. در واقع بقول اين محققين جنگي بين اسپرم هاي شامپانزه وجود دارد. هر شامپانزه نر سعي مي كند ژن هاي خود را تحميل نمايد. در واقع طي اين روند ، ژن هاي غير توليد مثلي واقع بر كروموزوم Y ، سست و غير فعال شده اند. بعلاوه تحليل Y در شامپانزه شباهت هايي را با انسان نشان داده است. دانشمندان اين مؤسسه بر اين باورند كه اگر چه كروموزوم Y انساني در ابتدا ژن هايي را از دست داده اند ، ولي در قرون اخير نسبتاً پايدار شده است.
كروموزوم Y انسان از سرنوشت Y شامپانزه دور مانده است و اين امر بدليل تك همسري بودن و يا منوگامي بودن انسان است.
اسپرم انسان رقابت اسپرم هاي شامپانزه را نداشته است. بنابراين فشاري را براي توليد اسپرم قوي تر متحمل نشده است. تمام ژنتيك دانان معتقدند كه زوال كروموزوم Y در انسان متوقف شده است. جني آ. مارشال از دانشگاه ملي استراليا ، كانبرا ، مي گويد كه از 300 مليون سال گذشته كه كروموزوم Y انسان متولد شد ، بتدريج تحليل رفته است. ولي همچنان بحث ادامه دارد كه آيا كاهش ميزان مردان در مقايسه با زنان متوقف شده است و يا خير ؟
|
منبع: Popular Science |
فاکتور X عامل افزایش طول عمر زنان
در مقایسه بین دو جنس زن و مرد ، طبیعت به زنان کمک بیشتری کرده است. 
طبق گفته متخصصان علم ژنتیک زنان دو کپی از کروموزوم قوی X را دارند در حالیکه مردان تنها با یک کروموزوم X و کروموزوم نسبتا ضعیف Y متولد می شوند.
به گزارش هلث دی تحقیقات ژنتیکی نشان میدهد علاوه بر مشخص کردن مرز DNA بین پسران و دختران ، این تقسیم به زنان توانایی بیشتری را در دفع بیماری می دهد. حتی در دروان جنینی نیز تعداد جنین های پسری که با ناهنجاری های ژنتیکی می میرند بیشتر از جنین های دختر است. آنها زودتر می میرند ، و مطمئنا آسیب پذیر ترند.
به گفته دکتر باربارا می گون استاد ژنتیک در دانشگاه جان هاپکینز آنچه در باره علل تفاوت های دو جنس می شنویم به تفاوت های مرد و زن در تجارب زندگی یا هورمون ها و مسائلی از این قبیل محدود می شود. و آنچه باقی مانده تفاوت ژنتیکی اساسی بین دو جنس است. این تفاوت از این واقعیت ناشی می شود که زنان با دو کپی از کروموزوم توانمند X متولد می شوند.
مردم فکر میکنند کروموزوم X فقط به جنسیت مربوط می شود اما این کروموزوم 1000 ژن دارد که در کارهای مختلف از جمله دخالت در انعقاد خون ، دخالت در عملکرد عضله و نیز در رها شدن از محصولات زائد سلولی دخالت دارند.
ژن های کروموزوم X می توانند جهش یابند یا دچار ناهنجاری شوند و این هم می تواند منجر به بیماری شود. بنابراین اگر یک جهش در ژنی روی کروموزوم X رخ دهد فردمذکر کپی سالم آن را ندارد و فقط گرفتار همین ژن معیوب خواهد بود. اما فرد مونث با داشتن دو کروموزوم X ممکن است یک کپی نرمال ژن را روی کروموزوم X دیگر داشته باشد.
این پدیده نشان می دهد که زنان در برابر بیمای های خاصی مقاوم اند. اما یک مشکلی وجود دارد که متخصصان ژنتیک آن را mosaicism می نامند. ویژگی فوق اشاره به این واقعیت دارد که بیان همزمان هر دو کپی از ژن های موجود روی کروموزوم X برای سلولها کشنده است. بنابراین در سلول های هر زن یکی از کروموزوم های X خاموش است در حالیکه دیگری فعال است.
اما بطور کلی این ویژگی یک مزیت محسوب می شود چرا که به سلولهای بدن زنان این امکلن را داده است در برابر جهش های خطرناک انعطاف پذیر تر باشند. اما در برخی موارد نادر این مزیت روند معکوس پیدا می کند. برای مثال پزشکان مدتهاست دریافته اند زنان بیشتر مستعد بیماری های اتوایمیون مانند ام.اس هستند.
به گفته دکتر می گون در اوائل دوران تکامل جنینی در صد کوچکی از زنان ممکن است متحمل عدم تعادل در بیان ژنتیکی کروموزوم X شوند که در آن یک طرف برنده می شود. این امر فرآیندهای مهمی مانند فرایند شناخت خودی (که در آن سیستم ایمنی یاد می گیرد سلولها و بافت های خودی را از غیر خودی تشخیص دهد) را تحت تاثیر قرار می دهد. اگر این فرآیند تشخیص بافت خودی از مسیر خود منحرف شود بیماری اتوایمیون رخ می دهد.
جام جم آنلاین
شواهد تازه از انتقال DNA بين حشرات خونخوار و پستانداران

دانشمندان در يك تحقيق جديد به شواهدي از انتقال مواد ژنتيكي بين حشرات خونخوار و پستانداران دست يافتهاند.
براساس اين گزارش محققان دانشگاه تگزاس در آرلينگتون امريكا به اولين مدارك و شواهد قطعي از انتقال افقي dna كه جابهجايي مواد ژنتيكي بين گونههاي بدون جفتگيري، بين بيمهرگان انگلي و برخي از ميزبانان مهرهدار آنها اطلاق ميشود، دست پيدا كردهاند.
محققان در اين پژوهش روي انتقال ترانسپوزون از نوعي حشره خونخوار بومي امريكاي جنوبي و يك حلزون آبگيري به ميزبانان آنها مطالعه كردند.
ترانسپوزون بخشي از مولكول وراثتي dna است كه ميتواند خود را تكثير كند و داخل ژنوم به موقعيتهاي مختلف تغيير مكان دهد.
اين فاكتور ميتواند سبب جهشهاي ژنتيكي، تغيير در ميزان dna در سلول و تأثير فاحش روي ساختار و عملكرد ژنوم در منطقه استقرار ترانسپوزون شود. اين تحقيقات نشان داد ترانسپوزونهايي كه در اين حشرات يافت ميشوند در 98 درصد ميزبانهاي پستاندار اين انگلها شناخته شده هستند.
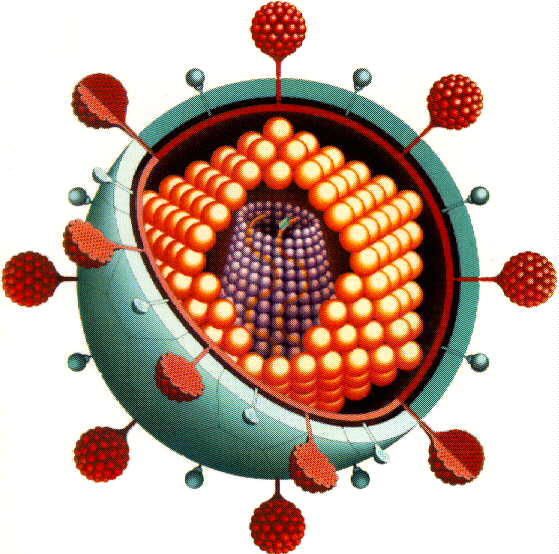
زیست شناسی ویروس ایدز (HIV)
اچ آی وی ويروسی است که به بيماری مرگبار ايدز منجر می شود. اين ويروس با حملات بی امان خود سيستم دفاعی بدن را چنان تضعيف می کند که حتی کوچکترين عفونت ها جان بيمار را به خطر می اندازند.

تاکنون دست کم 28 ميليون نفر در سراسر جهان در اثر ابتلا به ايدز در گذشته اند.
در حالی که 20 سالی است از شناسايی اچ آی وی می گذرد، هنوز هيچ واکسنی برای اچ آی وی و معالجه ای برای ايدز کشف نشده است. با اين حال نسل تازه داروها، طول عمر افراد آلوده به اچ آی وی را به طور چشمگيری افزايش می دهد.
ويروس اچ آی وی
اچ آی وی (Human Immunodeficiency Virus) به سيستم دفاعی، يعنی دقيقا همان سيستمی که معمولا در مقابل عفونت ها از بدن دفاع می کند، هجوم می برد.
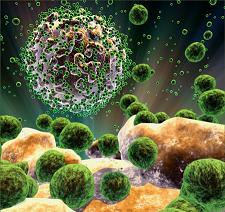
اين ويروس به يک نوع خاص از گلبولهای سفيد خون به نام "سی دی 4 +" حمله می کند. اچ آی وی اين سلول را ربوده، ژن های خود را وارد دی ان ای آن می کند، و از آن برای توليد ذرات ويروسی بيشتری استفاده می کند. اين ذرات سپس ساير سلول های سالم سی دی 4 + را آلوده می کنند.
آن دسته از سلول های سی دی 4 + که ميزبان ويروس اچ آی وی هستند در نهايت از بين می روند. نحوه نابودی اين سلول ها هنوز برای دانشمندان مجهول است.
با کاهش شمار سلول های سی دی 4 + توانايی بدن برای مبارزه با بيماری ها افت می کند و به تدريج به سطحی خطرناک می رسد. از اين مرحله به بعد بيمار به ايدز (Acquired Immune Deficiency Syndrome) مبتلا شده است.
اچ آی وی نوع خاصی از ويروسی به نام "رتروويروس" (retrovirus) است. هرچند اين رتروويرسها نسبت به ويروس های معمولی ارگانيسم ساده تری دارند، اما از ميان بردن آنها دشوارتر است.
ژن های رتروويروسها در داخل دی ان ای سلول ها لانه می کنند. به اين ترتيب هر سلول جديدی که از سلول ميزبان تکثير شود حاوی ژن های اين ويروس خواهد بود.
رتروويروس ها همچنين ژن های خود را با ضريب خطای بالا در سلول ميزبان کپی می کنند. توام شدن اين خصيصه با سرعت بالای تکثير اچ آی وی ، باعث می شود اين ويروس با همان سرعتی که منتشر می شود، تعيير شکل نيز بدهد.
علاوه بر اين، "پوششی" که ذرات ويروس اچ آی وی در آن محفوظ است، جنسی مشابه سلول های بدن دارد که باعث می شود سيستم دفاعی بدن نتواند ذرات ويروس را از سلول های سالم به راحتی تشخيص دهد.
شرح تصوير
اچ آی وی چگونه تکثير می شود:

1- ويروس خود را می چسباند: خوشه های پروتئينی در سطح بيرونی ذرات ويروس، به دريافت کننده های پروتئينی "سی دی 4 +" سلول مقصد "ملحق" می شوند.
2- ژن ها کپی می شوند: ويروس اچ آی وی نمونه ای از اطلاعات ژنتيکی خود را کپی می کند.
3- تکثير: ويروس اين کپی از ژن ها را وارد دی ان ای سلول ميزبان می کند. وقتی سلول ميزبان شروع به تکثير می کند، بخشهای تشکيل دهنده ويروس اچ آی وی را هم تکثير می کند.
4- ويروس جديد آزاد می شود: بخشهای تشکيل دهنده ويروس اچ آی وی در نزديکی جداره سلول جاسازی می شوند. آنها يک "جوانه" را تشکيل می دهند که از سلول جدا می شود و به يک ذره جديد اچ آی وی تبديل می شود.
آلودگی
ويروس اچ آی وی در خون، مايعات تناسلی افراد و همينطور شير مادر آلوده يافت می شود. اين ويروس همراه با اين مايعات وارد بدن ديگران می شود.
راه های سرايت اچ آی وی:

- آميزش جنسی با فرد آلوده بدون رعايت اقدامات پيشگيرانه
- استفاده اشتراکی از سرنگ آلوده يا ابزاری که برای سوراخ کردن اعضای بدن استفاده می شود و آلوده هستند
- از طريق خون آلوده
- در معرض تماس قرار دادن زخم يا بريدگی در بدن با مايعات آلوده به اچ آی وی
- نوزادان مادران آلوده ممکن است در دوران جنينی، هنگام تولد يا از طريق تغذيه از شير مادرآلوده شوند
ويروس اچ آی وی در بزاق دهان شخص آلوده وجود دارد، اما ميزان آن کم تر از حدی است که بتواند بيماری را منتقل کند. زمانی که مايعات حامل ويروس خشک شده باشد، خطر انتقال ويروس نزديک به صفر خواهد بود. برای پيشگيری از انتقال اچ آی وی، بهترين راه خودداری از برقراری رابطه جنسی با فرد آلوده به ايدز و استفاده از کاندوم های جنس "لاتکس" (نوعی پلاستيک) است.
ذرات با ابعاد اچ آی وی نمی توانند از کاندوم لاتکس عبور کنند و اگر به درستی و به طور منظم استفاده شوند شيوه کاملا موثری در کاهش خطر انتقال بيماری محسوب می شوند. هرچند تنها شيوه صد در صد موثر خودداری کامل از آميزش جنسی است.
معتادان تزريقی می توانند با خودداری از مصرف سرنگ های مشترک خطر آلودگی به اچ آی وی را کاهش دهند.
- از طريق هوا، سرفه و عطسه
- از طريق بوسيدن، تماس پوستی يا دست دادن
- از طريق استفاده مشترک از لوازم آشپزی مانند ديگ و چاقو
- از طريق تماس با صندلی توالت
- از طريق حشرات، نيش يا گاز حيوانات
- از طريق شنا در استخرهای عمومی
- از طريق خوردن غذايی که توسط فرد آلوده به اچ آی وی تهيه شده باشد
مراحل ابتدايی
حدود نيمی از افرادی که تازه به اچ آی وی آلوده می شوند در عرض دو تا چهار هفته به علائمی شبيه آنفولانزا مبتلا می شوند. اين علائم شامل تب، خستگی، ناراحتی پوستی، درد مفصل، سردرد و تورم در غدد لنفاوی است.

نمودار سمت چپ فرآيند تدريجی آلودگی به اچ آی وی را نشان می دهد. واحد شمارش گلبولهای سفيد سی دی 4 + برابر با تعداد آنها در يک ميليمتر مکعب خون است. با پيشرفت ويروس از تعداد آن کاسته می شود.
يک سيستم دفاعی سالم دارای 600 تا 1200 سلول سی دی 4 + در ميليمتر مکعب است. اگر اين رقم به 200 عدد کاهش يابد، بيمار مبتلا به ايدز محسوب می شود.
"بار ويروسی" شمار ذرات ويروسی در ميليمتر مکعب است. در مراحل ابتدايی، اين رقم با تکثير سريع ويروس در خون به اوج می رسد.
در برخی از افراد آلوده به اچ آی وی سالها طول می کشد تا بيماری ايدز ظاهر شود و در اين مدت آنها احساس سلامت کرده و هيچ علائم بيرونی حضور اين ويروس در آنها ديده نشود.
در ساير افراد آلوده به اچ آی وی ممکن است علائمی مانند کاهش وزن، تب و عرق، کهير و برای مدتی کوتاه پاک شدن حافظه نمود کند.
آزمايش های اچ ای وی:
- در جريان عمومی ترين آزمايش برای تشخيص اچ آی وی، يک نوع خاص از پادتن ها يا همان پروتئين های ضدبيماری رديابی می شود
- با اين که پادتن ها در واکنش به حضور اچ آی وی در بدن توليد می شوند، اما اين اتفاق شش تا دوازده هفته پس از آلودگی رخ می دهد
- در حالی که اين پادتن ها در مبارزه با ويروس چندان موثر نيستند، اما به عنوان نشانه وجود ويروس در بدن قابل اعتماد هستند
- در چند هفته ای که رديابی آلودگی بی ثمر و جواب آزمايش منفی است، فرد آلوده می تواند ويروس را به سايرين منتقل کند.
پيشروی بيماری ايدز در بدن

سيستم دفاعی بدن که از حمله ويروس صدمه ديده است، قوای خود را برای مبارزه با بيماری ها از دست می دهد و هر نوع عفونتی می تواند جان بيمار را تهديد کند.
کسانی که به اچ آی وی آلوده هستند در مقابل بيماری هايی مانند سل، مالاريا و ذات الريه آسيب پذيرتر می شوند و با افت شمار گلبولهای سفيد سی دی 4 +، آسيب پذيری آنها از قبل هم بيشتر می شود.
بيماران مبتلا به اچ آی وی همچنين در مقابل آنچه به "عفونت های فرصت طلب" معروف است آسيب پذير هستند. اين نوع از عفونتها از باکتری های شايع، قارچ ها و انگل هايی نشات می گيرد که يک بدن سالم قادر به مبارزه با آنهاست، اما می تواند در افرادی که سيستم دفاعی بدن آنها آسيب ديده ايجاد بيماری کند و گاه آنها را از پا در آورد.
برخی از اين عوامل بيماری زا زمانی که شمار سلول های سی دی 4 + زياد است به بدل حمله می کنند، اما اکثر آنها زمانی که شمار اين سلولها به کمتر از 200 رسيد، يعنی نقطه آغاز ظهور ايدز، فعال می شوند.
در صورتی که مراقبتهای کامل پزشکی در دسترس باشد، آن وقت می توان به بيمار داروهای درمانگر و محافظ در برابر برخی از اين عفونت ها را داد. البته اين داروها گاه گران است و به علاوه می تواند عوارض جانبی داشته باشد.
*برفک
برفک و زونا
برفک يک عفونت قارچی است که معمولا به دهان، گلو يا دهانه رحم حمله می کند.
ويروس زونا،"هرپيس سيمپلکس" (herpes simplex)، می تواند در دهان يا رحم ايجاد شود. هر دو عفونت شايع هستند، اما درصد وقوع آنها در ميان افراد مبتلا به اچ آی وی افزايش می يابد و حتی می تواند افرادی را که شمار سلول های سی دی 4 + آنها هنوز بالاست مبتلا کند.
علائم: برفک باعث پيدايش برآمدگی های سفيدرنگ در پوست و خشکی دهان می شود و فرآيند بلعيدن غذا را دشوار می سازد. زونا باعث پيدايش تاول های دردناک در ناحيه مبتلا می شود.
سل*
سل عامل اصلی مرگ و مير در ميان مبتلايان به ايدز در سراسر جهان است و بسياری از کشورها همزمان با اپيدمی های اچ آی وی و سل روبرو هستند. سل ناشی از يک نوع باکتری است که بسياری از مردم حامل آن هستند، اما تنها در برخی از افراد باعث بيماری می شود.
اگر افرادی که حامل اين باکتری هستند به اچ آی وی هم آلوده شوند، احتمال بروز سل در آنها 30 برابر افزايش می يابد. سل ابتدا به ريه حمله می کند، اما می تواند به غدد لنفاوی و مغز هم سرايت کند.
علائم: سرفه شديد، درد در قفسه سينه، سرفه همراه با خون، خستگی مفرط، کاهش وزن، تب و تعرق شديد در هنگام خواب.
سرطان سيستم دفاعی بدن*
افراد آلوده به اچ آی وی بيش از ديگران در معرض ابتلا به انواع سرطانهای سيستم دفاعی بدن موسوم به ان اچ ال (Non-Hodgkin’s Lymphomas) قرار دارند. ان اچ ال می تواند به هر بخشی از بدن از جمله نخاع و مغز حمله کند و می تواند ظرف يک سال بيمار را از پا درآورد. اين بيماری می تواند، صرف نظر از شمار گلبولهای سفيد سی دی 4 + در بدن بيمار اچ آی وی، بروز کند. شيمی درمانی راه مقابله با اين نوع سرطانهاست.
علائم (ان اچ ال): تورم غدد لنفاوی، تعرق به هنگام خواب و کاهش وزن.
شمار سی دی 4+ : زير 250
زخم های بدخيم (کی اس)
کی اس (Kaposi’s sarcoma) يک نوع بيماری مشابه سرطان بوده و در ميان مردان آلوده به اچ آی وی شايع است. اين بيماری موجب ظهور زخم های (ليژن) قرمز يا بنفش رنگی می شود که معمولا روی پوست ظاهر می شود. اين بيماری همچنين می تواند دهان، غدد لنفاوی، دستگاه گوارش و ريه ها را تحت تاثير قرار دهد و مرگبار باشد. اين بيماری معمولا به بيمارانی که شمار سی دی 4 + آنها کمتر از 250 است حمله می کند، اما در کسانی که شمار اين سلول در آنها کمتر است وخيم تر خواهد بود.
علائم: زخم، نفس تنگی در صورتی که به ريه حمله کرده باشد، و خونريزی در صورتی که به دستگاه گوارشی حمله کرده باشد.
شمار سی دی 4+ : زير 200
ذات الريه (پی سی پی)
موضع: ريه
پی سی پی (Pneumocystis pneumonia) يکی از عفونت های "فرصت طلب" است که باعث ذات الريه می شود. اين عفونت معمولا به ريه حمله می کند اما همچنين می تواند غدد لنفاوی، طحال، کبد يا مغز استخوان را هدف قرار دهد. اين بيماری همواره از علل عمده مرگ و مير در ميان بيماران آلوده به اچ آی وی بوده است، اما اکنون به کمک دارو قابل پيشگيری و معالجه است. اين بيماری اغلب افرادی را که شمار سی دی 4 + در آنها کمتر از 200 است مبتلا می کند.
علائم: تب، سرفه خشک، حس فشردگی در قفسه سينه و دشواری در عمل تنفس.
شمار سی دی 4+ : زير 100
عفونت های مغزی
بيماران اچ آی وی همچنين در مقابل دو نوع عفونت، که عموما به مغز حمله می کند، آسيب پذير هستند. توکسوپلاسموسيس (Toxoplasmosis) که از يک انگل در حيوانات ناشی می شود، می تواند باعث پيدايش زخم در مغز شود. کريپتوکوکوس (Cryptococcus) - نوعی قارچ در خاک - اغلب باعث مننژيت می شود. اين بيماری در پرده نخاع و مغز عفونت ايجاد می کند و می تواند به اغما و مرگ منجر شود. اين عفونت ها در بيمارانی که شمار سلول های سی دی 4 + در آنها کمتر از 100 است شايع است.
علائم: سردرد، تب، مشکلات بينايی، تهوع و استفراغ، ضعف در يک طرف بدن، دشوار شدن تکلم و راه رفتن (توکسوپلاسموسيس)، گرفتگی عضلات گردن (مننژيت).
شمار سی دی 4+ : زير 75
عفونت شکم (ام ای سی)
ام ای سی يا ام ای آی (Mycobacterium avium complex) عفونتی است که از باکتری های موجود در آب، گرد و غبار، خاک و مدفوع پرندگان ناشی می شود. اين بيماری به لايه بيرونی معده و روده حمله می کند و در نهايت می تواند در خون و ساير بخش های بدن منتشر شود. ام ای سی اغلب کسانی را که شمار گلبولهای سفيد سی دی 4 + در آنها کمتر از 75 است مبتلا می کند.
علائم: شکم درد ، تهوع و استفراغ که به تب می انجامد، تعرق در خواب، بی اشتهايی، کاهش وزن، خستگی، اسهال.
شمار سی دی 4+ : زير 50
خطر نابينايی (سی ام وی)
سی ام وی (Cytomegalovirus) يک عفونت مرتبط با ويروس زونا (هرپيس) است و در بيماران اچ آی وی اغلب باعث ابتلا به تورم شبکيه چشم (retinitis) می شود. اين بيماری با مرگ سلول های شبکيه در ناحيه عقب چشم همراه است و در صورتی که معالجه نشود به سرعت می تواند باعث نابينايی شود. سی ام وی توسط دارو قابل مهار است. اين ويروس همچنين می تواند بر ساير بخش های بدن اثر بگذارد. اين بيماری به ندرت به افرادی که شمار سلول های سی دی 4 + آنها بيش از 100 است حمله می کند، و اغلب افرادی را که شمار اين سلول در آنها کمتر از 50 است مبتلا می کند.
علائم: مشکل بينايی مانند ديدن لکه های سياه متحرک، تاری در ديد و پيدايش نقاط کور.
داروهای ضد ايدز
از زمان آغاز اپيدمی اچ آی وی، مجموعه ای از داروها ساخته شده اند که از طريق مهار توانايی تکثير اين ويروس، عمر افراد مبتلا به اچ آی وی را به طور قابل ملاحظه ای افزايش می دهد. اين داروها از سرعت نابودی سی دی 4 + بيماران می کاهد و می تواند بروز ايدز را به تاخير بياندازد، اما آن را معالجه نمی کند.
در مجموع چهار دسته دارو وجود دارد که در مقاطع مختلف آلودگی و رشد اچ آی وی و ايدز عمل می کنند:

1) بازدارنده های ورودی: اين داروها به پروتئين های موجود بر سطح بيرونی ويروس اچ آی وی می چسبد و از پيوستن و ورود آن به سلول های سی دی 4 + جلوگيری می کند. تاکنون تنها يک نمونه از دارو، به نام "فوزيون" (Fuzeon)، وارد بازار شده است.
2) بازدارنده های ان آر تی (Nucleoside reverse transcriptase): اين بازدارنده ها ويروس اچ آی وی را از نسخه سازی از ژنهای خود باز می دارد. نوکليوسايدها مصالح ساختمانی اين ژنها هستند. اين دارو فرآيند نسخه سازی را از طريق توليد نمونه های معيوبی از اين مصالح ساختمانی مختل می کند.
3) بازدارنده های ان ان آر تی (Non-nucleoside reverse transcriptase): اين بازدارنده ها نيز فرآيند نسخه سازی را مختل می کنند. آنها با چسباندن خود به آنزيمی که اين فرآيند را کنترل می کند نسخه سازی را مختل می کنند.
4) بازدارنده های نوع پروتيز (Protease): اين داروها به آنزيم ديگری به نام پروتيز که نقشی اساسی در جمع آوری ذرات ويروس تازه دارد، می چسبند.
داروهای ضدويروس ايدز بايد به صورت ترکيبی مصرف شوند. معمولا سه نوع داروی مختلف از دست کم دو دسته مختلف از داروها به طور همزمان به بيمار تجويز می شود. با تغيير شکل دادن ويروس اچ آی وی، برخی از نمونه های اين ويروس در مقابل داروها مقاوم می شوند. بنابراين شانس کنترل اچ آی وی در صورت استفاده از چند دارو بيشتر خواهد بود. در برخی موارد از آلودگی های تازه، ويروسهايی رديابی شده اند که حتی پيش از آغاز معالجات در برابر داروهای موجود مقاومت نشان می دهند.
عوارض جانبی
عوارض جانبی شايع:
- تهوع، استفراغ، سردرد، خستگی مفرط، کهير، اسهال، بی خوابی، بی حسی در اطراف دهان، درد معده
ساير عوارض جانبی
- التهاب لوزالمعده، آسيب به کبد و لوزالعمده، زخمهای درون دهان، تغيير شکل بدن، آسيب به سلولهای عصبی، کم خونی، درد عضلانی و ضعف.
ویروس ایدز (HIV) ویروسی است که باعث نقص ایمنی بدن میشود و تولید بیماری میکند. ویروس ایدز (HIV) مخفف (Human Immuno deficincy Virus) است. این ویروس در گروه رترو ویروسها قرار دارد.
اطلاعات تکبیلی دیگر راجع به ویروس ایدز:
ویروس ایدز (HIV) ویروسی است که باعث نقص ایمنی بدن میشود و تولید بیماری میکند. ویروس ایدز (HIV) مخفف (Human Immuno deficincy Virus) است. این ویروس در گروه رترو ویروسها قرار دارد.
رترو ویروسها ، حاوی ژنوم RNA همراه با RNA پلیمراز وابسته به DNA (ترانس کریپتاز معکوس) میباشند. ژنوم رترو ویروس از دو زیر واحد مشابه ، هر یک حاوی RNA تک رشتهای مثبت ، به اندازه 7 تا 11 هزار کیلو باز تشکیل شده است. ذرات ویروسی حاوی ترانس کریپتاز معکوس هستند که برای همانند سازی ویروس ضروری است. ذرات ویروس حاوی ریبو نوکلئو پروتئین مارپیچی بوده و درون یک کپسید 20وجهی قرار دارند.
سه گروه مورفولوژیک از ذرات رترو ویروس شناخته شدهاند. ویروسهای نوع سوم بزرگترین رترو ویروسها هستند که بعضی ویروسها جزء این گروه میباشند. یکی از زیر خانوادههای رترو ویروسها ، لنتی ویرینه است که شامل عواملی هستند که قادرند عفونتهای مزمن با تخریب آهسته و پیشرونده عصبی ایجاد کنند. ویروس نقص ایمنی انسان در این گروه قرار دارد.
خصوصیات مهم لنتی ویروسها
-
این ویروسها دارای پوشش هستند.
-
گلیکو پروتئینهای پوشش از لحاظ آنتی ژنی تغییر میکنند.
-
برای همانندسازی آنزیم ترانس کریپتاز معکوس ، نسخهای از ژنوم DNA را از روی RNA ژنومی تهیه میکند. DNA پرو ویروس به عنوان الگویی برای RNA ویروسی عمل میکند، تغییرات ژنتیکی شایع است.
-
اعضای آن سرطانزا نیستند و سلولهای دستگاه ایمنی را آلوده میکنند. پرو ویروسها به صورت مادامالعمر در سلول باقی میمانند. بیان ویروس در بعضی سلولهای بدن محدود میباشد، بیماری مزمن آهسته و پیشروندهای ایجاد میکنند. همانند سازی به میزان زیادی اختصاصی است. گروه شامل عامل ایجاد کننده AIDS است.
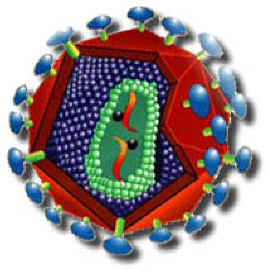
ساختمان و ترکیب ویروس HIV
این موفقیت مرهون ویرولوژی مولکولی پیشرفته است که تنها 4 سال پس از شناخته شدن یک بیماری و سندرم غیر معمول به نام ADIS ، در سال 1981 ، عامل ایجاد کننده آن شناسایی و جداسازی شده است. HIV یک رترویروس و عضو زیرخانواده لنتی ویرینه است و بسیاری از خصوصیات نمادین فیزیکی _ شیمیایی خانواده خود را داراست.
خصوصیت مورفولوژیک منحصر به فرد HIV ، وجود نوکلوئیدی استوانهای در ویریون بالغ است. این نوکلوئید میلهای شکل که دارای ارزش تشخیصی میباشد، در تصاویر میکروسکوپ الکترونی دیده میشود. این ویروس سه ژن لازم برای همانند سازی را دارا میباشد. حدود 6 ژن اضافی دیگر ، بیان ویروس را تنظیم میکنند و در پاتوژنز بیماری در بدن حائز اهمیت هستند. نمونههای جدا شده مختلف HIV مشابه نیستند، اما ظاهرا طیفی از ویروسهای مرتبط به هم را تشکیل میدهند. گروههای مختلفی از ژنوم ویروسی در افراد آلوده یافت شده است.
طبقه بندی
لنتی ویروسها از گونههای بسیاری از جمله 20 گونه مختلف از پریماتها (نخستیها مانند میمونها) جدا شدهاند. دو نوع متمایز از ویروسهای انسانی ADIS به نام HIV-1 و HIV-2 وجود دارند. افتراق این دو نوع ویروس از طریق ساختار ژنومی و ارتباط فیلوژنیک با سایر لنتی ویروسهای پریمات ، صورت میگیرد. 
منشا ADIS
اعتقاد بر این است که HIV انسانی از عفونتهای بین گونهای توسط ویروسهای میمون در مناطق روستایی آفریقا ، نشات گرفته است. احتمالا انتقال توسط تماس مستقیم انسان با خون پریماتهای آلوده صورت گرفته است و بوجود آمدن تغییرات خاص اجتماعی ، اقتصادی در ابتدا و اواسط قرن 20 زمینه را برای گسترش عفونت ویروسی ، تثبیت عفونت آن در انسان و ایجاد اپیدمی فراگیر ، فراهم ساخته است.
ضد عفونی کردن و غیر فعال سازی ویروس
HIV در دمای اتاق در عرض 10 دقیقه توسط هر یک از مواد زیر کاملا غیر فعال میشود:
-
محلول سفید کننده خانگی کلردار 10 درصد
-
اتانول 50 درصد
-
ایزوپروپانول 35 درصد
-
پراکسید هیدروژن 0.3 درصد
همچنین ویروس در دو انتهای طیف PH غیر فعال میشود، اما هنگامی که HIV در خون لخته شده یا نشده ، در سرنگ یا سوزن وجود داشته باشد، باید حداقل 30 ثانیه در معرض محلول سفید کننده کلر غیر رقیق قرار گیرد، تا غیر فعال شود. ویروس توسط tween-20 ، دو و نیم درصد غیر فعال نمیشود. گر چه پارافورمالدئید ویروس آزاد در محلول را غیرفعال میکند، مشخص نیست که آیا به اندازه کافی در بافت نفوذ میکند تا تمام ویروسهای موجود در سلولهای کشت داده شده و یا نمونههای بافتی را غیرفعال سازد یا خیر؟
سیستمهای لنتی ویروس حیوانی
الگوی بیماری طبیعی در گونههای مختلف متفاوت است، اما تعدادی خصوصیات مشترک شناخته شدهاند:
-
ویروسها از طریق تبادل مایعات بدن انتقال مییابند.
-
ویروس بطور نامشخص در میزبان آلوده باقی میماند. هر چند ممکن است در سطوح بسیار کم حضور داشته باشد.
-
میزان جهشها بالا است و انتخاب جهشهای مختلف ، بستگی به شرایط متفاوتی دارد که ویروس در آن به سر میبرد (فاکتورهای میزبان ، پاسخ ایمنی و نوع بافتها). میزبانهای آلوده محتوی گروههایی از ژنوم ویروسی موسوم به شبه گونه میباشند که رابطه بسیار نزدیکی با یکدیگر دارند.
-
روند عفونت ویروسی به آهستگی طی مراحل مخصوص پیشرفت میکند. رده سلولی ماکروفاژها نقشی محوری در عفونت ایفا میکنند. لنتی ویروسها از این نظر که میتوانند سلولهای انتهایی تمایز یافتهای را که قابلیت تقسیم ندارند، آلوده کنند با سایر رترو ویروسها متفاوت هستند.
-
ممکن است تا ایجاد بیماری سالها طول بکشد. میزبانان آلوده معمولا بر ضد ویروس آنتی بادی تولید میکنند، اما قادر به پاک سازی عفونت نمیباشند. در نتیجه ویروس برای تمام عمر با میزبان باقی میماند.
-
علائم بالینی میتواند از 3 ماه تا چندین سال پس از عفونت ، در هر زمانی ایجاد شود. بطور استثنا در برخی بیماریهای لنتی ویروس دوره کمون طولانی نمیباشد. از جمله ADIS در کودکان ، آنمی عفونی در اسبها.
فاکتورهای میزبانی موثر در پاتوژنز
-
سن: افراد جوان در خطر بیشتری هستند.
-
استرس: استرس ممکن است سبب آغاز بیماری شود.
-
ژنتیک: بعضی از گونههای حیوانات مستعدتر هستند.
-
عفونتهای همزمان: ممکن است سبب تشدید بیماری یا تسهیل انتقال ویروس شود.
گیرندههای ویروسی
تمامی لنتی ویروسهای پریمات از مولکول 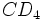 به عنوان گیرنده استفاده میکنند. این مولکول در سطح ماکروفاژها و لنفوسیتهای T وجود دارد. برای ورود HIV به داخل سلول علاوه بر به گیرنده دیگری نیاز است. گیرنده دوم برای ادغام ویروس با غشای سلول لازم میباشد. گیرندههای کموکین به عنوان گیرنده دوم HIV عمل میکنند.
به عنوان گیرنده استفاده میکنند. این مولکول در سطح ماکروفاژها و لنفوسیتهای T وجود دارد. برای ورود HIV به داخل سلول علاوه بر به گیرنده دیگری نیاز است. گیرنده دوم برای ادغام ویروس با غشای سلول لازم میباشد. گیرندههای کموکین به عنوان گیرنده دوم HIV عمل میکنند.
کموکینها
فاکتورهایی محلول هستند که خاصیت سیتوکینی و جذب شیمیایی دارند.
گسترش جهانی ADIS
بر اساس آمار سال 2000 ، بیشترین میزان عفونت در نواحی زیر صحرای آفریقا بوده است. در برخی شهرهای آفریقا که شیوع عفونت بالاست حدود بک سوم از بالغین به ویروس آلوده هستند. عفونت به سرعت در حال گسترش به جنوب شرقی آسیا است. به این دلیل که ایدز غالبا بالغین جوان و نیروی کار جامعه را مبتلا میسازد، اپیدمی ایدز اثرات زیانباری بر ساختار اجتماعی و اقتصادی کشورها بر جای گذاشته است.
تصور بر این است که انتشار سریع و جهانی ایدز در اواخر قرن 20 از مهاجرت عظیم جوامع روستایی به مراکز شهری همراه با نقل و انتقال افراد آلوده در نواحی مختلف جهان به دلیل معضلات شهری ، گردشگری و مسافرتهای شغلی ، نشات گرفته است.
مقدمه ای بر ویروس شناسی
ویروس شناسی (Virology) شاخهای از میکروبیولوژی (Microbiology) میباشد. این علم به بررسی ویروسها ، شکل و ساختمان آنها ، ژنتیک ویروسها و سایر خصوصیات آنها میپردازد.
دید کلی
تا اواخر قرن 19 اصطلاح ویروسهای پالایهپذیر برای مشخص کردن عوامل عفونی که از صافیهای عبور دهنده باکتریها ، قارچها و پروتوزوئرها میگذرند، اطلاق میشد. چند سال بعد اصطلاح پالایهپذیر حذف شد و کلمه ویروس به معنی سم اختصاصا به عوامل عفونی پالایهپذیر غیر قابل رویت با میکروسکوپ نوری اطلاق گردید. طی دهه اول قرن 20 اغلب دانشمندان عقیده داشتند که ویروسها عوامل عفونی مشخصی هستند که فقط از نظر اندازه با سایر میکروارگانیسمها تفاوت دارند. ولی به زودی کشف گردید که ویروسها روش خاص خود را برای تولید مثل دارند و ترکیب شیمیایی آنها مشخص میباشد. ابداع میکروسکوپ الکترونی و پیشرفت در روشهای تجزیهای موجب پیشرفت در شناسایی ساختمان و اختصاصات فیزیولوژیکی ویروسها شد. ویروسها به چند دلیل مورد توجه خاص میکروبیولوژیستها قرار دارند.
-
این ذرات اساسا با کلیه میکروبهایی که شناخته شده، از نظر ساختمان و چرخه زندگی تفاوت دارند.
-
گرچه برای اغلب عفونتهای باکتریایی داروهای متعددی در دسترس قرار دارد ولی برای بیشتر عفونتهای ویروسی داروی موثری وجود ندارد و از اینرو ویروسها در اغلب کشورها بیماریهای تهدید کننده حیات انسان ایجاد میکنند.
-
به احتمال زیاد ، ویروسها با برخی از انواع سرطانها در انسان رابطه دارند.

تاریخچه
علم ویروس شناسی ، پس از استفاده از واکسن آبله توسط ادوارد بوخنر (1798) ، تهیه واکسن علیه بیماری هاری بوسیله پاستور (1884) و انتقال بیماری موزائیک توتون از گیاه آلوده به گیاه سالم توسط مایر در سال 1886 آغاز شده است. در سال (1892) ایوانوسکی ، ثابت کرد که عامل مولد بیماری موزائیک توتون از صافیهای بسیار ریز نگهدارنده باکتری عبور میکند. ولی نتوانست به اهمیت مساله پی ببرد. تا این که بیچرنیک (1898) دانشمند هلندی که باید او را پدر ویروس شناسی به حساب آورد ادعا کرد که عامل بیماری موزائیک توتون ، باکتری نیست و عامل مسری دیگری است. استنلی در سال 1935 با استفاده از روشهای جدید خالص کردن پروتئینها ، ویروس را خالص کرد و به پاس این خدمت ، موفق به دریافت جایزه نوبل شد.
صفات عمومی ویروسها
ویروسها عواملی هستند که واجد یک نوع اسید نوکلئیک هستند. دارای پوشش پروتئینی در اطراف اسید نوکلئیک میباشند. در درون سلولهای زنده با بکار گرفتن ماشین سنتزی سلول میزبان تکثیر پیدا میکنند و اسید نوکلئیک ویروسی را به سایر سلولها منتقل میسازند. چون ویروسها فاقد آنزیمهای لازم برای متابولیسم هستند، لذا برای تکثیر یافتن بایستی از ماشین متابولیکی سلول میزبان استفاده نمایند. این کیفیت از نظر پزشکی جهت پیدا کردن داروهای ضد ویروسی فوقالعاده اهمیت دارد. زیرا اکثر داروهایی که تکثیر ویروسها را متوقف میسازند، برای اعمال سلول میزبان نیز اثر میگذارند. ولی وجود لیپیدها در سطح خارجی برخی از ویروسها آنها را در برابر حلالهای لیپیدها نظیر اتر ، آسیبپذیر میسازد.
اساس رده بندی ویروسها
میزان اطلاعات قابل دسترس در هر زمینه برای تمامی ویروسها ، یکسان نیست و روشی که براساس آن ویروسها شناسایی میشوند، به سرعت در حال تغییر است. امروزه اکثرا از بررسی توالی ژنی به عنوان یک روش اولیه برای شناسایی ویروس استفاده میشود و بدین ترتیب نیاز به سایر اطلاعات کلاسیک نظیر چگالی شناوری ویروس ، کاهش یافته است. دادههای مربوط به توالی ژنی معیارهای پیشرفته طبقه بندی محسوب شده و گاهی باعث ایجاد خانوادههای جدیدی از ویروسها میشوند. مورفولوژی ویروس ، خصوصیات فیزیکی- شیمیایی ویروس ، خصوصیات ژنومی ویروس ، خصوصیات بیولوژیک و ... در طبقه بندی ویروسها مد نظر قرار میگیرند.

روشهای انتقال و انتشار ویروسها
ویروسهای مختلف دارای مکانیزمهایی دقیق و پیچیده برای ادامه حیات و انتقال از یک میزبان به میزبانی دیگر هستند. روش انتقال یک ویروس از یک میزبان به میزبان دیگر به ماهیت واکنش متقابل بین ویروس و میزبان بستگی دارد. ویروسها به روشهایی انتقال مییابند مثلا انتقال از فردی به فرد دیگر در اثر تماس مستقیم. بیشترین راههای انتقال ویروسها توسط قطرات تنفسی ، ذرات عفونی معلق در هوا و تماس جنسی است. ویروسهای گیاهی
بیشتر از طریق بند پایان منتقل میشوند. منشا تکاملی ویروسها
انواع مختلف ویروسها از نظر منشا تکاملی با یکدیگر تفاوت دارند. دو تئوری مطرح شده در این مورد به شرح زیر است:
تئوری اول
منشا ویروسها ممکن است از RNA ، DNA و یا از هر دو نوع اسید نوکلئیک سلولهای میزبان باشد که بطور خودمختار همانند سازی کرده و روند تکاملی خود را طی نمودهاند. و در واقع ویروسها شبیه ژنهایی هستند که توانایی موجودیت مستقل از سلول را کسب کردهاند. توالی ژنی در برخی از ویروسها با ژنهای سلولی رمز کننده پروتئینهای عملکردی قرابت دارند. به نظر میرسد که حداقل برخی از ویروسها بدین روش ، تکامل یافتهاند.
تئوری دوم
ویروسها ممکن است اشکال انحطاط یافته انگلهای داخل سلولی باشند. هیچ گونه مدرکی در دست نیست که نشان دهد ویروسها از باکتریها تکامل یافته اند. اما ممکن است منشا تکاملی ارگانیسمهای داخل سلولی اجباری نظیر ریکتسیاها و کلامیدیاها ، مربوط به باکتری باشد. به هر حال پولکس ویروسها به قدری بزرگ و پیچیده هستند که احتمالا از تکامل بعضی اجداد سلولی بوجود آمدهاند.

تکثیر ویروسها
اسید نوکلئیک هر ویروس فقط تعداد معدودی از ژنهای لازم برای سنتز ویروسهای جدید را دارا میباشد. این ژنها شامل ژنهای سازنده اجزای سازنده آنزیمهای لازم در چرخه زندگی ویروس میباشد. برای تکثیر ویروس بایستی ویروس سلول میزبان را مورد حمله قرار داده و اختیار دستگاه متابولیکی آن را به عهده گیرد. در جریان تکثیر ویروسی ، یک ذره ویروسی ، صدها حتی هزاران ویروس تولید میکند. این تغییرات شدید در سلول میزبان ، به مرگ آن منجر میگردد.
ارتباط ویروس شناسی با سایر علوم
ویروس شناسی با رشتههای میکروبیولوژی ، زیست شناسی سلولی ، ژنتیک ، انگل شناسی ، قارچ شناسی پزشکی ، بیوشیمی ارتباط نزدیک دارد
مقدمه ای بر ژنتیک جمعیت
جمعیت از نظر ژنتیکی عبارت است از گروهی از موجودات یک گونه که با یکدیگر آمیزش پیدا میکنند. گروهی محدود از جمعیت که با هم ، آمیزش دارند، ژنتیک مندلی هم گفته میشوند. ژنتیک جمعیت ، شاخهای از علم ژنتیک است که رفتار ، خصوصیات ، فراوانی و عمل متقابل ژنها را در یک جمعیت مندلی که دارای ذخایر ژنی مشترک هستند، بطور ریاضی بر اساس قانون تعادل هاردی _ وینبرگ ، مورد تجزیه و تحلیل قرار میدهد.
اختلاف نظر بین صاحبنظران در مورد نقش ژنتیک در تکامل موجودات با پیدایش علم سنتتیک جدیدی به نام ژنتیک جمعیت (Population genetics) در دهه 1920 از بین رفت و متعاقبا دانشمندان مختلف از جمله هاردی ، ریاضیدان انگلیسی و وینبرگ در سال 1908 مطالعه نحوه رفتار ژنها و تغییرات فراوانی آنها در جمعیت و نقش آنها در تکامل موجودات زنده را دنبال و مهمترین قانون مرتبط با ژنتیک جمعیت را زیر عنوان قانون هاردی _ وینبرگ در اوایل این قرن پیشنهاد کردند. بطوری که به کمک این قانون میتوان بسیاری از جنبههای مختلف ژنتیک جمعیت را مورد بحث قرار دارد.
سرنوشت یک جفت ژن در یک جمعیت به چه صورت است؟ قدرت تولید مثل یک موجود که دارای ژن بخصوصی است، بستگی به فراوانی آن ژن در جمعیت ، و عواملی دیگری از جمله رابطه بین آن جمعیت و محیط دارد. بنابراین هر چند افراد حامل ژن میباشند، ولی سرنوشت این افراد و ژنی را که حمل میکنند، بستگی به جمعیت و عوامل موثر در آن دارد. جمعیت چه هاپلوئید و چه دیپلوئید و ... باشد، دارای دو صفت ویژه است: فراوانی ژنی و حوضچه ژنتیکی.
-
فراوانی ژنی عبارت است از نسبت آللهای مختلف یک ژن در جمعیت. جهت بدست آوردن فراوانی ژنی ، تعداد افرادی را که دارای ژنوتیپهای مختلف هستند، بدست آورده و نسبت فراوانی نسبی هر کدام از آللها را تخمین میزنیم. فراوانی یک ژن از فراوانی ژنوتیپ هموزیگوس نسبت به آن ژن به اضافه نصف فراوانی هتروزیگوسها هم محاسبه میشود.
-
حوضچه ژنتیکی عبارت است از مجموع ژنهای موجود در گامتهای تولید شده توسط یک جمعیت. بنابراین رابطه ژنتیکی بین یک نسل با نسل دیگر شبیه رابطه ژنتیکی بین والد و نوزاد است.

جمله بینومی  و ژنتیک جمعیت
و ژنتیک جمعیت
در این جمله حروف a نماینده احتمال حدوث یک اتفاق ، b نماینده احتمال حدوث اتفاق دیگر و n نماینده تعداد اتفاقات است. جمله را میتوان برای بیان نسبت ژنوتیپی 1:2:1 حاصل از تلاقیهای مونوهیبریدی بکار برد. به جای a از حروف p و به جای b از حروف q استفاده میشود. از آنجایی که در یک مکان ژنی معمولا دو تا آلل وجود دارد، فراوانی آنها را در جمعیت مجموعا برابر واحد یک در نظر میگیرند. با توجه به اطلاعات فوق میتوان ژنوتیپهای موجود در یک مونوهیبرید ( ) را به صورت معادله
) را به صورت معادله 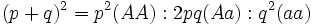 ، بیان نمود.
، بیان نمود.
محاسبه فراوانی اللها
هم بارزی یا غالبیت ناقص
در جمعیتهای انسانی دو تا الل اتوزومی M ( ) و N (
) و N ( ) وجود دارند که میتوانند آنتی ژن خون و نوع آن را تحت تاثیر قرار دارند. در جهت تعیین فراوانی اللهای و مطالعهای در مورد مهاجرین قفقازی کشور ایالات متحده به صورت زیر گزارش شده است:
) وجود دارند که میتوانند آنتی ژن خون و نوع آن را تحت تاثیر قرار دارند. در جهت تعیین فراوانی اللهای و مطالعهای در مورد مهاجرین قفقازی کشور ایالات متحده به صورت زیر گزارش شده است:
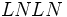
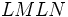
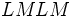 ژنوتیپ
ژنوتیپ 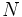
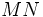
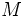 فنوتیپ 130330391787تعداد
فنوتیپ 130330391787تعداد
برای محاسبه فراوانی اللهای و میتوان به صورت زیر عمل کرد:


غالبیت کامل
غالب و مغلوب بودن اللها ، فراوانی آنها را مستقیما تحت تاثیر قرار نمیدهد. فراوانی آنها مشابه اللهای هم بارز یا اللهای با غالبیت ناقص است. الل غالب در مقایسه با الل مغلوب ، فنوتیپ مربوطه را در توده بیشتر ظاهر میسازد، چرا که الل غالب اثر الل مغلوب را در حالت هتروزیگوسی نیز نهفته نگه میدارد. قدرت چشایی و عدم قدرت چشایی نسبت به نمک فنیل تیو کاربامید (PTC) مثال مناسبی برای مطالعه فراوانی اللها در حالت غالبیت کامل است. کسانی که دارای قدرت چشایی نسبت به این نمک هستند، مزه آن را تلخ احساس میکنند و این صفت تحت کنترل یک ژن غالب T قرار دارد.
چند اللی
در صورتی که یک ژن بیش از دو آلل داشته باشد، با استفاده از بسط دو جملهای میتوان فراوانیهای ژنوتیپی هر کدام از ژنوتیپها را بدست آورد. برای مثال اگر ژنی دارای سه آلل  ،
، 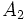 ،
، 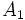 با فراوانیهای به ترتیب p ، q و r باشد، بطوری که
با فراوانیهای به ترتیب p ، q و r باشد، بطوری که 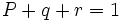 ، فراوانی ژنوتیپی را در حالت تعادل میتوان از بسط سه جملهای زیر بدست آورد که فراوانیهای ژنوتیپی در گروههای خونی ABO را در انسان میتوان با این روش نشان داد.
، فراوانی ژنوتیپی را در حالت تعادل میتوان از بسط سه جملهای زیر بدست آورد که فراوانیهای ژنوتیپی در گروههای خونی ABO را در انسان میتوان با این روش نشان داد.
OOBB BOABAA AOژنوتیپ OBABAفنوتیپ 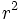


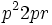
فراوانی وابستگی به جنس
در مورد ژنهای وابسته به جنس ، تعداد ژنوتیپهای ممکنه افزایش مییابد. علت این افزایش تفاوت کروموزومهای جنسی در جنسهای نر و ماده است. اگر مادهها XX و نرها XY باشند، از نظر یک جفت ژن a و A پنج ژنوتیپ امکانپذیر است، سه عدد از این ژنوتیپها (AA , Aa , aa) در مادهها و دو عدد در نرها (A , a) وجود خواهد داشت. اگر آمیزش به صورت تصادفی باشد، میتوان ثابت کرد که این تعادل نسل به نسل ، باقی خواهد ماند. این تعادل بر مبنای برابر بودن فراوانی آللها در نر و ماده است. اگر تفاوتی بین فراوانی اللها در دو جنس مخالف وجود داشته باشد، جمعیت در حالت تعادل نمیباشد.

برای مونثها
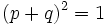

آمیزشهای خویشاوندی
از انواع آمیزشهای غیر تصادفی ، آمیزشهای خویشاوندی یا همخونی است. در این نوع آمیزش ، افرادی که دارای قرابت و خویشاوندی هستند، با هم آمیزش پیدا میکنند. همخونی دارای درجات مختلفی است. نزدیکترین نوع آن خودلقاحی است که در گیاهان صورت میگیرد. گیاهانی که نسبت به یک جفت ژن هتروزیگوس هستند، در اثر خودلقاحی تولید نوزادانی میکنند که 50% آنها هموزیگوت و مابقی هتروزیگوت هستند. میزان افزایش هموزیگوسیتی در اثر ازدواجهای فامیلی را ضریب همخونی Coefficient inbreeding گویند و آن را با f نشان میدهند.
ضریب همخونی احتمال به ارث رسیدن دو آلل یک جایگاه ژنی در یک موجود است که منشا مشترک داشته باشند و یا به عبارت دیگر این آللها کپی یک ژن در یک والد مشترک باشد. این ضریب در مورد فرد بکار برده شده و درجه خویشاوندی بین والدین فرد را نشان میدهد. اگر دو والد به صورت تصادفی آمیزش پیدا کنند، ضریب همخونی نوزاد برابر این است که دو گامت به صورت تصادفی از والدین دارای ژنهای یکسان در یک جایگاه ژنی باشند.
اصل هاردی _ وینبرگ
این اصل بدین صورت بیان میگردد که در یک جمعیت بزرگ که آمیزش به صورت تصادفی است و مهاجرت ، جهش و انتخاب وجود ندارد، فراوانی ژنی و ژنوتیپی ، نسل به نسل ، ثابت باقی مانده و فراوانی ژنوتیپی را میتوان به کمک فراوانی ژنی بدست آورد.
عواملی که فراوانی ژنها را تغییر میدهند
-
موتاسیون: جهش موجب پیدایش اللهای جدید و نهایتا تغییرات ژنتیکی میشود.
-
مهاجرت: حرکت افراد یا ژنها از یک جمعیت به جمعیت دیگر را در ژنتیک ، مهاجرت گویند.
-
رانش ژنتیکی: در جمعیتهای کوچک ، فراوانی پارهای از آللها ، ممکن است بطور تصادفی شدیدا تغییر یابد.
-
آمیزشهای غیر تصادفی: خویش آمیزی باعث میشود که فراوانی پارهای از ژنوتیپها از آنچه که قانون هاروی _ وینبرگ پیش بینی میکند، متفاوت باشد.
-
گزینش: گزینش با تغییر در فراوانی ژنها ، عامل مهم در تغییرات تکاملی در داخل یک جمعیت به شمار میرود و میتواند موجب جدا شدن یا تفکیک جمعیتها به نژادها و گونههای مختلف شود.
تعادل ژنتیکی
ژنتیک به علت تاکید آن بر خود بیمار و نیز خانواده او ، در بین شاخههای گوناگون پزشکی ، منحصر به فرد است. ژنتیک جمعیت ، مطالعه توزیع ژنها در جمعیت و چگونگی حفظ یا تغییر فراوانی ژنها و ژنوتیپها میباشد. ژنتیک جمعیت با عوامل ژنتیکی مانند جهش ، تولید مثلبیماریهای ژنتیکی در خانوادهها و جوامع را تعیین میکنند.
اصل زمینهای ژنتیک جمعیت ، قانون تعادل هاردی - وینبرگ است. لازم است بتوانیم با استفاده از ارقام مربوط به بروز یک بیماری ارثی یا منفعت ژنتیکی دیگر ، فراوانی ژنوتیپهای خاص را تعیین کنیم و از آنها فراوانی آللهای خاص مسئول ژنوتیپهای مختلف را استنباط کنیم. اگر جمعیتی برخی از خصوصیات را داشته باشد یک رابطه ساده ریاضی به نام قانون هاردی - وینبرگ ، برای محاسبه ژنوتیپها از روی فراوانی آللهای وجود دارد.
و نیز با عوامل محیطی و اجتماعی مانند انتخاب و مهاجرت سر و کار دارد که اینها همراه با یکدیگر فراوانی و توزیع خصوصیات یک جمعت متعادل :
-
تمام افراد جمعیت بایستی از نظر تولید مثل باهم یکسان باشند.
-
جمعیت باید از تعداد زیادی افراد تشکیل شده باشد.
-
آمیزش به صورت تصادفی باشد.
-
نباید مهاجرت به درون یا به بیرون جمعیت صورت بگیرد.
-
بایستی جهش در حالت تعادل باشد.
نقص هر کدام از شروط بالا میتواند باعث تغییر فراوانی ژنها و لذا بهم خوردن تعادل ژنتیکی گردد.
خصوصیات قانون تعادل
خاصیت اول قانون تعادل
فراوانی سه ژنوتیپ AA (غالب) Aa ، aa (مغلوب) ، به صورت بسط دو جملهای p+q)2 = p2 + q2 + 2pq) نشان داده میشود. p فراوانی آلل A و q فراوانی آلل a در استخر ژنی است و ترکیب آللها به صورت تصادفی بوده و ژنوتیپهای حاصل تصادفی هستند. احتمال ایجاد ژنوتیپ AA برابر p2 ، aa برابر q2 و Aa برابر 2pq است.
خاصیت دوم قانون
درصد ژنوتیپها از نسلی به نسل بعد تغییر نمیکند. یعنی هرگاه در جمعیتی که ژنوتیپهای AA , Aa , aa با نسبتهای p2 : 2pq : q2 وجود دارند، لقاح تصادفی صورت بگیرد، فروانی ژنوتیپها در نسل بعضی به صورت درصدهای نسبی ثابت خواهد ماند. معادله هاردی - وینبرگ هیچ مقدار خاصی را برای q , p مشخص نمیکند. فراوانی آللها در جمعیت به هر مقداری که باشد موجب فراوانی ژنوتیپی به صورت p2 : 2pq : q2 میشود، مادام که فراوانی آلل ثابت بماند، این فراوانیهای ژنوتیپی از نسلی به نسل دیگر ثابت خواهند بود.
نتایج قانون تعادل
اگر توان دوم فراوانی ژنوتیپی هتروزیگوتها در یک جمعیت برابر چهار برابر حاصل ضرب فراوانی دو ژنوتیپ هموزیگوت ، در جمعیت باشد، در این حالت میگوییم که جمعیتها به صورت متعادل است. در یک جمعیت بزرگ که آمیزشها به صورت تصادفی انجام میشود، و تمام ژنوتیپها از نظر قدرت زنده ماندن یکسان هستند. فراوانی ژنی در یک نسل بستگی به فروانی ژنی و نه فروانی ژنوتیپی نسل قبل دارد. جمعیتی که دارای این خاصیت باشد، اصطلاحا گویند در حالت تعادل (equilibrium) است. طبق قانون هاردی - وینبرگ اگر یک جمعیت در حال تعادل نباشد. فقط یک نسل آمیزش ، کافی است که آن را به حالت تعادل در آورد. 
کاربرد قانون تعادل
کاربرد عملی اصلی قانون هاردی - وینبرگ در ژنتیک پزشکی در مشاوره ژنتیکی برای اختلالات اتوزومی مغلوب است. در مورد بیماری فنیل کتونوریا ، فروانی هموزیگوتهای مبتلا در جمعیت را میتوان دقیقا تعیین کرد، زیرا بیماری از طریق برنامههای غربالگری در نوزادان شناسایی میشود. افراد هتروزیگوت ناقلان خاموش هستند و اندازه گیری مستقیم میزان بروز آنها در جمعیت از روی فنوتیپها غیر ممکن است. قانون تعادل هاردی - وینبرگ ، برآورد فراوانی هتروزیگوتها و استفاده از آن برای مشاوره و ژنتیکی را مقدور میسازد.
عواملی که تعادل هاردی - وینبرگ را بهم میزنند
در دنیای واقعی ژنتیک پزشکی شامل جمعیتهای انسانی و آللهای بیماری ، خصوصیات جامعه متعادل صدق نمیکنند ژنوتیپهای موجود در یک جمعیت ممکن است در تعادل هاردی - وینبرگ نباشند. از عوامل بر هم زننده تعادل میتوان آمیزشهای غیر تصادم را نام برد که شامل آمیزشهای خویشاوندی و غیر خویشاوندی است. از طرف دیگر بعضی باعث تغییر فراوانی ژنهای میشوند که شامل موارد زیر هستند.
مهاجرت
میزان تغییر فراوانی ژنی در جمعیتی که مهاجرت در آن صورت میگیرد بستگی به میزان مهاجرت و تفاوت بین فراوانیهای ژنومی بومیها و مهاجریسن دارد.
جهش
اگر ژن جدیدی در اثر جهش در یک جمعیت بوجود آید احتمال بقای این ژن بسیار کم است ولی عاملی که باعث تغییر فراوانی در اثر جهش میشود، فراوان بودن جهش است.
گزینش
یکی از مهمترین عوامل تغییرات فراوانی ژنها قدرت باروری حاملین آن است. انواع مکانیزمهایی که قدرت باروری یک ژنوتیپ را تغییر میدهد، گزینش نام دارد.
رانش یا دریفت
رانش ژنتیکی به علت اینکه جوامع از نظر اندازه محدود هستند، ایجاد میگردد. بطوری که خطای نمونهای سبب تغییرات فراوانی آنی میگردد.
چشم انداز بحث
توصیف ریاضی رفتار ژنها در جمعیتها با جز مهمی در بسیاری از شاخهها مانند انسان شناسی ، زیست شناسی تکاملی و ژنتیک انسانی است. در ژنتیک پزشکی ، ژنتیک جمعیت عمدتا کاربرد عملی داشته است. یعنی تعیین فراوانی آللی محاسبات خطر بیماری. با افزایش نقش آزمایشهای ژنتیکی و فناوریهای درمانی در بزرگتر جلوه دادن اثر مراقبتی ژنتیک پزشکی بر سلامت عمومی و خطر بیماریهای ژنتیکی در نسلهای بعد درک اصول ژنتیک جمعیت روز به روز مهمتر خواهد شد.
تئوری تکامل
موجودات زمین در طول زمین طی فرآیندی مستمر ، تحول پیدا میکنند که تکامل نامیده میشود. از زمانی که زندگی بر روی زمین آغاز شد، میلیونها گونه جدید پدید آمده و از بین رفتهاند. موجودات زنده امروزی ، تنها نمایانگر بخش کوچکی از تمام موجودات زندهای هستند که تا به حال وجود داشتهاند. بطور عمده تکامل در نتیجه انتخاب طبیعی رخ میدهد. این فرآیند در جهت حفظ بقا است. بنابراین به قدرت تولید مثل افرادی که سازگاری بیشتری نسبت به دیگر اعضا همان گونه با محیط دارند بستگی دارد.
بین افراد هر گونه با گونههای دیگر تفاوتهایی وجود دارد. ممکن است بعضی گونهها به خاطر احتیاج به غذای کمتر و یا دوری از حیوانات شکاری نسبت به گونههای دیگر شرایط بهتری را دارا باشند. این افراد احتمالا بهتر میتوانند به بقای خود ادامه داده و ساختار بدنی مطلوب خود را به نسلهای بعدی خود منتقل کنند. این فرآیند ، که به نام انتخاب طبیعی معروف است، گونهها را قادر میسازد تا خود را با محیط متغیر سازگاری داده و در دراز مدت منجر به پدید آمدن گونههای جدید میشوند.

قدیمیترین نوشتهای که به بحث درباره آفرینش جانداران پرداخته در تورات آمده است. بر اساس این نوشته خداوند جهان و جانداران ساکن آن را در شش روز آفریده و آدمی آخرین آفریده او است. نظریههای بعدی عبارت بودند از خلقالساعه و ثبوت انواع که تا قرن 18 و 19 تائید میشدند. فرانچسکو ردی پزشک ایتالیایی قرن 17 نخستین کسی بود که مدارکی علیه نظریه خلقالساعه بدست آورد. وی با استفاده از آزمایش نشان داد که موجود زنده نمیتواند از مواد بیجان پدید آید. در قرن 19 اعتقاد به خلقالساعه با کمک آزمایشهای پاستور روی باکتریها از اعتبار افتاد.
نظریه لامارک در مورد تکامل
در قرن 19 اندیشه پیوستگی جانداران و پیوستگی تاریخی آنها یعنی نطریه تکامل به فکر معدودی از متفکران رسیده بود. یکی از تئوریهای مهم تکاملی از لامارک زیست شناس فرانسوی است که به سال 1809 به چاپ رسید. لامارک برای بیان چگونگی وقوع تکامل دو نظریه استعمال و عدم استعمال اندامها و ارثی بودن صفات اکتسابی را عنوان کرد. وی مشاهده کرده بود که اگر اندامی از بدن یک جاندار استعمال میشود بزرگتر و کارآمدتر میشود و اگر عضوی بکار نیفتد کوچک میشود و تحلیل میرود.
بنابراین جاندار در نتیجه ناهماهنگی در استعمال و عدم استعمال اندامهای مختلف بدن در طول عمر خود ممکن است تا حدی تغییر یابد و بعضی از صفات را کسب کند. لامارک این گونه صفات را وراثتی و قابل انتقال به اخلاف میپنداشت. این تئوری بسیار موفقیت آمیز بود و به اشاعه اندیشه تکامل کمک بسیار کرد. اما سرانجام معلوم شد که نظر لامارک نادرست است. چون هر گونه تغییری که بر اثر استعمال یا عدم استعمال یا به هر علت دیگر در سلولهای دیگر بدن به غیر از گامتها رخ دهد تاثیری در ژنهای سلولهای زاینده نخواهد داشت و قابل انتقال به نسل بعدی نیست. 
در سال 1858 داروین و والاس باهم تئوری جدیدی درباره تکامل اعلام داشتند که جانشین تئوری لامارک شد. داروین این تئوری جدید را پروراند و به صورت کتابی تنظیم کرد. این اثر معروف که به نام اصل انواع بوسیله انتخاب طبیعی یا بقای نژادهای مناسب در تنازع بقا است در سال 1859 انتشار یافت. داروین و والاس محیط را علت اصلی انتخاب طبیعی میدانستند، یعنی محیط تدریجا جانداران دارای صفات نامساعد را از بین میبرد و جانداران دارای صفات مساعد را حفظ میکند. پس از گذشت نسلهای زیاد و متوالی و تاثیر مداوم انتخاب طبیعی ، سرانجام گروهی جاندار یک صفت یا تعدادی صفات جدید و مساعد را به درجهای جمع خواهد کرد که به صورت گونهای جدید از گونه اجدادی ظاهر خواهد شد.
همواره این سوال مطرح میشد که منبع این تفاوتهای فردی مهم چیست؟ و تفاوتهای فردی چگونه بوجود میآیند. اینجا بود که داروین ناگزیر شد به نظریه لامارک یعنی وراثت صفات اکتسابی پناه ببرد. اما تدوین پاسخ درست مربوط به تفاوتهای فردی شش سال پس از انتشار کتاب اصل انواع و هنگامی آغاز شد که مندل قوانین وارثت را اعلام داشت. ولی حاصل مطالعات مندل بیش از 30 سال در ابهام نادیده ماند و پیشرفت شناخت مکانیسمهای تکاملی نیز به همان نسبت به تاخیر افتاد. انتخاب طبیعی فقط به عنوان بخشی از مکانیزم تکامل شناخته شده است. زیرا داروین مانند لامارک در شناسایی علل ژنتیکی تغییرات تکاملی توفیق بدست نیاورده بود. داروین و والاس توضیحی ناقص از تکامل عرضه داشتند، اما بطور کلی آنان نخستین کسانی بودند که جهت درست تکامل را نشان دادند.
گوناگونی ژنتیکی
مکانیزم تکامل را میتوان اثر انتخاب طبیعی بر تفاوتهای ژنتیکی که در میان افراد یک جمعیت ظاهر میشوند، دانست. جمعیت ، گروهی از افراد یک نوع جاندار است که در یک محل زندگی کرده و باهم زاد و ولد میکنند و ژنها آزادانه در میان آنها توزیع میشوند. گوناگونی ژنتیکی ممکن است از نوترکیبی حاصل از تولید مثل جنسی ، از جهش یا از هر دو این پدیدهها ناشی نشود. اگر چنین جاندارانی باقی بمانند و اولاد بیاورند، در آن صورت خاصههای ژنتیکی جدید آنها در خزانه ژن جمعیت باقی خواهد ماند. این تازههای ژنتیکی در طول نسلهای متوالی میتوانند در تعداد بیشتر ی از افراد جمعیت یا در همه آنها گسترش یابند.
مفهوم واقعی انتخاب طبیعی ، تولید مثل افتراقی است. یعنی بعضی از افراد یک جمعیت بیش از دیگر افراد آن اولاد به جا میگذارند و نسبت به افرادی که اولاد کمتری از آنها میماند، درصد بیشتری از ژن به خزانه ژنی نسل بعد ، انتقال میدهند. اگر تولید مثل افتراقی در طول چند نسل ادامه یابد، آنهایی که فرزندان بیشتری تولید میکنند، تدریجا سهم بزرگتری از افراد کل جمعیت را بوجود خواهند آورد. در نتیجه ژنهای آنها در خزانه فزونتر خواهد شد. پس امکان دارد صفت جدیدی که در یک فرد جاندار پدید میآید، از راه تولید مثل افتراقی و به مرور زمان ، به صورت خصوصیت استاندارد تمامی یک جمعیت درآید.
نتیجهگیری کلی
بر اساس نظریه امروزی تکامل ، انتخاب طبیعی نیرو در اساس خلاقه به شمار میآید که نوآورهای ژنتیکی را در میان افراد بیشتری گسترش میدهد. هرچند که به از میان رفتن افرادی که از نظر تولید مثل صلاحیت ندارند، میانجامد. افرادی را که از نظر رفتاری ، یا از نظر اجتماعی ، صلاحیت ندارند، الزاما از میان نمیبرد. قویترین و بزرگترین جاندار یک جمعیت ممکن است از نظر رفتاری بسیار صالح باشند، ولی اگر نازا باشند، از نظر تولید مثل و از نظر تکامل ، فاقد صلاحیت خواهند بود.
از سوی دیگر ممکن است فردی رنجور و ضعیف ، از نظر رفتاری یا اجتماعی صلاحیت نداشته باشد، اما اولاد زیادی بوجود آورد. انتخاب طبیعی در اساس از طریق تولید مثل واقع میشود نه از راه تنازع بقا. تنازع بقا برای بقا روی میدهد، ولی غالبا جنبه فیزیکی محض دارد و ممکن است به صورتی غیر مستقیم بر موفقیت جاندار در امر تولید مثل نیز موثر افتد. این عوامل نیز در حد خود میتوانند اثرات تکاملی به بار آورند. اما مهمترین نتیجه حاصل نه تنازع ، نه از میان رفتن و نه حتی بقای فرد است. بلکه آنچه سرانجام حاصل میشود، موفقیت نسبی در امر تولید مثل است.
ژن های مرتبط با آلزایمر کشف شدند
کشف دو ژن اولین سرنخی است که در طی شانزده سال اخیر در حوزه تحقیقات در باره آلزایمر به دست آمده و باعث شده است محققان نظریه های خود را در باره چگونگی به وجود آمدن این بیماری در انسان بازنگری کنند.
این ژن ها در جریان مطالعه شانزده هزار نمونه از دی ان ای رد یابی شده است و سابق بر این محققان آنها را در ایجاد تورم در داخل بدن انسان و همچنین تجزیه کلسترول موثر می دانسته اند.
این امید وجود دارد که مطالعه ای که نتیجه آن در نشریه علمی "نیچر ژنتیک" منتشر شده است راه را برای درمان های جدید هموار کند.
تنها ژنی که با شکل متدوال تر بیماری آلزایمر مرتبط دانسته شده، ژن APOE4 است که محور اغلب تحقیقاتی بوده که تا کنون انجام گرفته است.
اطلاعات به دست آمده از تازه ترین تحقیق، که کار گروهی دانشگاه های بریتانیا بوده است، در اختیار محققان فرانسوی که سومین ژن، ژن CR1، را شناسایی کرده اند، گذاشته شده است.
دو ژنی که توسط گروه بریتانیایی کشف شده اند نقش حفاظتی در مغز ایفا می کنند.
تغییر در این ژن ها ممکن است تاثیر حفاظتی آنها را ضایع کند یا آنها را از نقش "محافظ به مهاجم" تبدیل کند.
پرفسور کوین مورگان، یکی از اعضای این گروه تحقیقی از دانشگاه ناتینگهام، می گوید فعل و انفعالاتی که به خاطر این کشف جدید مشخص شده، فعل و انفعالاتی است که در تجزیه کلسترول و همچنین بخشی از دستگاه ایمنی، که در ایجاد تورم در داخل بدن انسان دست دارد، نقش دارد.
او گفت: "فعل و انفعالاتی که جدیدا کشف شده اند، راه های تازه ای را برای درمان دارویی آلزایمر باز می کند."
آقای مورگان گفت: "اکنون سوال این است که آیا با کاهش کلسترول بد در خون و همچنین کاهش تورم این امکان وجود دارد که خطر ابتلا به آلزایمر کمتر شود یا نه."
جولی ویلیامز، که هدایت تحقیقات جدید را برعهده داشته است گفت: "ما به نظریه های خاصی در باره بیماری آلزایمر توجه داشته ایم اما اطلاعات ما نشان می دهد که اتفاقات مختلفی در بدن انسان موجب این بیماری می شود."
او گفت: "ما واقعا نمی دانیم چه چیزی باعث این بیماری می شود. در چند سال آینده ما ممکن است درک خوبی از این بیماری داشته باشیم."
شمار بیماران مبتلا به بیماری آلزایمر در سراسر جهان رو به افزایش بوده است و فقط در بریتانیا 700 هزار نفر به این بیماری، که علاوه بر اخلال در حافظه فرد باعث معلولیت های جسمی و ذهنی زیادی نیز می شود، مبتلا هستند.انتظار می رود تا سال 2050 تعداد مبتلایان به یک میلیون و هفتصد هزار نفر افزایش یابد.
منبع: BBC
هوش در گیاهان
یکی از تفاوتهای آشکار بین ما جانوران و خویشاوندان سبز رنگ دورمان ، یعنی گیاهان ، میزان جنبش و جابهجایی ماست. ما پذیرفتهایم که هوش را از روی کارها بسنجیم، زیرا کارهایی که انجام میدهیم نشان میدهند که در مغز ما چه میگذرد. بنابراین، چون گیاهان خاموش و بیجنبش به چشم میآیند و در یک جا ریشه دواندهاند، زیاد تیز هوش و زرنگ به نظر نمیرسند. اما گیاهان نیز جنبش دارند و به برانگیزانندههای پیرامون خود پاسخ میدهند.
گیاهان با حساسیت چشمگیری دستکم 15 متغیر محیطی گوناگون را پیوسته بررسی میکنند. آنها میتوانند این پیامهای ورودی را پردازش کنند و با کمک دستهای از مولکولها و راههای پیامرسانی ، خود را برای پاسخ درست آماده سازند. بنابراین ، توان محاسبهگری گیاهان بیمغز شاید به اندازه بسیاری از جانوران بامغزی باشد که میشناسیم. ساقه در حال رشد میتواند با کمک پرتوهای قرمز دور(مادن قرمز) ، نزدیکترین همسایههای رقیب خود را حس کند و پیامد کارهای آنها را پیشبینی کند و اگر لازم باشد، به شیوهای از رخدادن آن پیامدها پیشگیری کند.
برای مثال ، هنگامی که همسایههای رقیب به نخل (Stilt) نزدیک میشوند همه گیاه به سادگی جابهجا میشود. ریزوم برخی گیاهان علفی با رشد کردن به سوی بخش بدون رقیب و یا سرشار از مواد غذایی ، جای زندگی خود را بر میگزیند. سس که نوعی گیاه انگل است، طی یک یا دو ساعت پس از نخستین برخوردش با گیاه میزبان ، توانایی بهرهبرداری از آن را میسنجد. خلاص ه، گیاهان میتوانند ببینند، بچشند، لمس کنند، بشنوند و ببویند.
دوری از سایه
بخش دریافتکننده نور ساختمان تتراپیرولی دارد و از راه اسید آمینه سیستئین به بخش دگرگونکننده که گونهای پروتئین است، پیوند میشود. فیتوکروم در پاسخ به طول موجهای گوناگون نور به شکل کارا و ناکارا درمیآید. شکل ناکارا (Pr) پس از جذب فوتونهای قرمز به شکل کارا (Pfr) درمیآید. Pfr که فوتونهای قرمز دور (مادون قرمز) را بهتر دریافت میکند، در پاسخ به این طول موجها به Pr دگرگون میشود.
پس از درک نسبت پایینی از نور قرمز به قرمز دور ، گیاهی که از سایه دوری میگزیند (گیاه آفتاب پسند) بر رشد طولی خود میافزاید و اگر ترفنندهایش کارگر افتند، جنبههای دیگر پاسخ دوری از سایه باعث شتاب گرفتن گلدهی و تولید پیش از زمان دانه میشوند تا بخت ماندگاری افزایش یابد. دانشمندان در آزمایشی گروهی از گیاهان را زیر فیلتری پرورش دادند که نسبت نور قرمز به قرمز دور را کاهش میداد و بنابراین ، پاسخ دوری از سایه را برمیانگیخت. این گیاهان نسبت به گیاهانی که زیر نور کامل خورشید میروییدند، رشد طولی بیشتری پیدا کردند. البته ، اندازه رشد طولی به اندازه آفتابپسندی گیاه ارتباط دارد. گیاهان صحرایی نسبت به گیاهانی که بطور معمول در سایهی درختان چنگل میرونید، رشد طولی بیشتری پیدا کردند.
فیتوکرومها اغلب فعالیت پروتئینکینازی را از خود نشان میدهند. این مولکولها با پیوند زدن گروههای فسفات به پروتئینها ، فعالیت آنها را تغییر میدهند. بر این اساس ، آنها با تغییر فعالیت پروتئینهایی که در تنظیم ژنها دخالت دارند، بر فعالیت آنها تاثیر میگذارند. ژنهای زیادی در گیاهان شناخته شدهاند که از راه فیتوکروم در پاسخ به نور تنظیم میشوند. البته ، فیتوکرومها بخشی از پاسخهای زیستی را از راه تغییرهایی در تعادل یونها در سلول پدید میآورند.
هنگامی که همسایگان نخل پایهدار بر میزان نور دریافتی گیاه تاثیر میگذارند یا به منبع غذایی آن دست درازی میکنند، نخل فرار را برقرار ترجیح میدهد و همه گیاه به جایی جابهجا میشود که بسیار آفتابی است. برای این جابهجایی ریشههای تکیهگاهی جدید به سوی جای آفتابی رشد میکنند و ریشههای طرف سایهانداز شده آرامآرام میمیرند. در این رفتار گیاه ، به خوبی هدفدار کار کردن را میبینیم.
این گیاه همانطور که روی زمین میخزد، در دو بعد رشد میکند. هر جا که مناسب باشد، از ساقه زیر زمینی آن ریشههایی به سوی زمین و ساقههایی به سوی بالا پدید میآیند. وقتی گیاه در خاک مرغوبی قرار گیرد، انشعاب و شاخ و برگ بیشتری تولید می کند. همچنین ، تودههایی از ریشه پدید میآورد تا با سرعت بیشتری از خاک قطعهای که در آن میروید، بهرهبرداری کند. اما هنگامی که این گیاه خزنده در قطعهی فقیرتری قرار میگیرد، با سرعت بیشتری گسترش خود را به بیرون از آن قطعه ، پیش میبرد تا به هر گونهای از آن جا فرار کند.
در این حالت ، ساقهی زیر زمینی گیاه نازکتر است و انعشاب کمتری دارد. این تغییر در الگوی رشد باعث میشود، ساقههای هوایی جدید دورتر از گیاه والد شکل گیرند و در محیط تازهای به جست و جوی مواد غذایی بپردازند. البته ، میزان رشد فقط با کیفیت مطلق یک قطعه ارتباط ندارد، بلکه میزان مرغوبیت آن در مقایسه با قطعههای پیراون نیز برای گیاه مهم است. در واقع ، گیاه قطعهای را به عنوان قطعه مرغوب شناسایی میکند که دست کم دو برابر سرشارتر از قطعههای پیرامون باشد. اما پیش از این پاسخهای هوشمندانه ، گیاه باید بتواند کیفیت قطعهای را که در آن میروید بسنجد.
دو پژوهشگر انگلیسی ژنی را در گیاه رشادی (Arabidopsis) کشف کردهاند که به ریشهها این توانایی را میدهند که برای پیدا کردن قطعههای سرشار از نیترات و نمکهای آمونیوم ، خاک را بچشد. فراورده این ژن به ریشهها امکان میدهد به جای جست و جوی تصادفی و پر هزینه ، به سوی مواد غذایی رشد کنند. این دو پژوهشگر برای شناسایی ژنهایی که ممکن است در این کار دخالت داشته باشند، جهش یافتههای گوناگونی از رشادی را پرورش دادند تا سرانجام جهش یافتهای را پیدا کردند که نمیتوانست با توسعه ریشههای جانبی از ریشههای اصلی ، به جست وجوی نیترات بپردازد. به این ترتیب آنان ژنی را کشف کردند که برای شناسایی نیترات ضروری است.
گیاهان به کمک این آنزیم بخشی از فسفات معدنی لازم برای رشد خود را بدست میآورند. این دو پژوهشگر در آزمایشی نشان دادند، گیاهان تراژنی که مقدار زیادی آپیراز تولید میکردند، نسبت به گیاهان دیگر، رشد بیش تری داشتند. مکندههای گیاه سس (Cuscuta) نیز برای غارت بهترین گیاه میزبان از حس چشایی بهره میگیرند. این گیاه که توان فتوسنتز کردن ندارد، به گرد ساقههای میزبان می پیچد و برای بدست آوردن مواد غذایی و آب ، ساختارهای مکنده خود را درون آنها فرو میکند.
هوش این انگل گیاهی در ارزیابی مقدار انرژی که میتوان از میزبان بدست آورد و مقدار انرژی که برای بهرهبرداری از آن باید صرف شود، به کمک گیاه میآید. از لحظه برخورد انگل با گیاه میزبان تا آغاز گردآوری مواد غذایی از آن ، نزدیک 4 روز است. این زمان برای ارزیابی میزان پرباری میزبان و تصمیم گیری برای تولید پیچهای کم تر یا بیشتر به دور آن ، کافی است. پیچهای بیشتر به تولید مکندههای بیشتر و در نتیجه بهرهبرداری بیشتر از میزبان میانجامند. اما اگر میزبان پربار نباشد تولید پیچهای بیشتر نوعی هدر دادن انرژی به شمار میآید.
در دهه 1990 کولین کلی نشان داد راهبردهایی که گیاه سس برای جست و جوی بهترین میزبان بکار میگیرد، با مدلهای ریاضی که برای توضیح جنبههای اقتصادی جست و جوی غذا در جانوران ابداع شده بودند، هماهنگی دارند. بنابراین ، سس ممکن است زرنگترین شکارچی پیرامون ما نباشد، اما در جست و جوی شکار به خوبی جانورانی که میشناسیم، کار میکند.
از اینرو ، گیاهان میکوشند با تقویت بافتهای بخشهایی که به نوسان درمیآیند، در برابر باد پایداری کنند. البته ، هزینه کردن انرژی برای بافتها ممکن است کشاورزان را نگران کند. در یک آزمایش مشاهده شد وقتی گیاه ذرت هر روز به مدت 30 ثانیه تکان داده شود، میزان محصول تا 30 الی 40 درصد کاهش مییابد. پژوهشگران میخواهند بدانند چگونه پیام لمس ، بافتهای محکمتری تولید میکند. بیشتر پژوهشهای کنونی روی کلسیم متمرکز شده است. هنگامی که گیاهان به سویی کشیده میشوند، یونهای کلسیم از واکوئلها به درون سیتوزول جریان پیدا میکنند.
بیرون رفتن این یونها ، که تنها یک دهم ثانیه به درازا میکشد، به فعال شدن ژنهایی میانجامد که با تقویت دیواره سلول ارتباط دارند. تاکنون پنج ژن از این ژنهای لامسه (TCH) شناسایی شدهاند. یکی از این ژنها، رمز ساختن پروتئین کالمودولین را در خود دارد که حسگر اصلی کلسیم در گیاهان و جانوران است. در سال 1995 جانت برام چهارمین ژن لامسه (TCH4) را کشف کرد که آنزیمی به نام زیلوگلوکان اندوترانس گیکوزیلاز را رمز میدهد. این آنزیم روی دیواره سلولی گیاهان اثر میگذارد و با تغییرهایی که در اجزای اصلی سازنده آنها پدید میآورد، بر قدرت و استحکام آنها میافزاید.
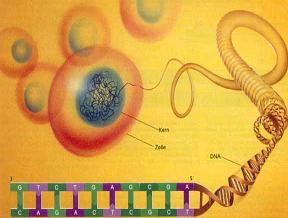
مقدمه ای بر ساختمان DNA
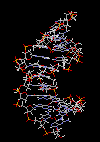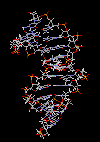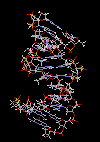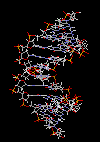
سرعت پیشرفت تعیین ساختمان DNA بسیار کند بوده است. در سال 1930 کاسل و لویننوکلئوتید میباشد که از سه قسمت تشکل شده است. یک قند پنتوز (2- دزوکسی D- ریبوز) ، یک گروه 5-فسفات و از یکی چهار باز آلی نیتروژندار حلقوی آدنین (A) ، گوانین (G) ، سیتوزین (C) و تیمین (T) تشکیل شده است.
از این چهار باز دو باز آدنین و گوانین از دریافتند که نوکلئین در واقع اسید دزوکسی ریبونوکلئیک است. برسیهای شیمیایی آن مشخص کرد که زیر واحد تکرار شونده اصلی DNA ، بازهای پورینی و دو باز سیتوزین و تیمین از بازهای پیریمیدینی میباشند. به مجموعه قند و باز آلی نوکلئوزید گفته میشود. گروه فسفات میتواند به کربن3 و یا5 متصل شود. به مجموع نوکلئوزید و گروه فسفات متصل به آن نوکلئوتید میگویند. با توجه به اینکه یون فسفات میتواند هم به کربن 3 و هم به کربن5 متصل شود.
پس دو نوکلئوتید از طریق یک پیوند فسفودی استر بهم متصل میشوند. به این صورت که گروه هیدروکسیل یک نوکلئوتید با گروه فسفات نوکلئوتید دیگر واکنش داده و پیوند فسفودی استر را بوجود میآورد. از آنجایی که پیوند فسفودی استر ، کربنهای3 و5 دو قند مجاور را بهم متصل میکند، این پیوند را پیوند5-3 فسفودی استر نیز مینامند. یک زنجیره در اثر اتصال پشت سر هم تعدادی2-دزوکسی ریبونوکلئوتید بوسیله پیوندهای دزوکسی ریبونوکلئوتید تشکیل میشود.
تمامی نوکلئوتیدها در یک زنجیره پلی نوکلئوتیدی دارای جهت یکسان میباشند. به این صورت که نوکلئوتید انتهایی در یک سمت زنجیره دارای یک گروه5 آزاد و نوکلئوتید انتهایی در سمت دیگر زنجیره دارای یک گروه3 آزاد میباشد. بنابراین زنجیره پلی نوکلئوتیدی دارای جهت بوده و این جهت را به صورت5--->3 نشان میدهند. بنابراین اگر در نوکلئوتید ابتدایی کربن5 در بالای حلقه پنتوز و کربن3 در زیر آن باشد، در تمامی نوکلئوتیدهای بعدی زنجیره کربن 5 در بالای حلقه پنتوز جای خواهد داشت.
-
DNA یک پلیمر رشتهای متشکل از واحدهای2- دزوکسی اسید ریبونوکلئیک میباشد که بوسیله پیوندهای فسفودی استر5-3 به هم متصل شدهاند.
-
DNA حاوی چهار زیر واحد dc و dG و dT و dA میباشد.
-
مقادیر متوالی dT و dA با یکدیگر و dc و dG نیز با یکدیگر مساوی میباشند.
مارپیچ دو رشتهای DNA
در سال 1953 در ساختمان سه بعدی DNA ، بوسیله واتسون و کریک کشف شد. واتسون و کریک با استفاده از مطالعات تفرق اشعه ایکس ، رشتههای DNA که بوسیله فرانکلین و ویلکینز تهیه شده بود و همچنین ساختن مدلها و استنباطهای مشخصی ، مدل فضایی خود را ارائه دادند و در سال 1962 واتسون و کریک و ویلکینز به خاطر اهمیت کشف ساختمان DNA به صورت مشترک جایزه نوبل دریافت کردند.
مدل پیشنهادی آنان چنین بود. DNA یک مارپیچ دو رشتهای است که رشتههای آن به دور یک محور مرکزی ، معمولا به صورت راست گرد پیچ میخورند. طبق مدل واتسون و کریک ، ستونهای قند - فسفات همانند نردههای پلکان به دو قسمت خارجی بازهای آلی پیچیده و به این ترتیب در معرض محیط آبکی داخل سلول هستند و بازهای آلی که خاصیت آبگریزی دارند، در داخل مارپیچ قرار میگیرند. هنگام تشکیل مارپیچ رشتهها به صورت موازی متقابل قرار میگیرند.
یعنی اگر جهت یک رشته3<--5 باشد، رشته دیگر 5<--3 خواهد بود. پیوندهای هیدروژنی بین آدنین از یک رشته با باز تیمین رشته مقابل و باز گوانین یک رشته با سیتوزین رشته مقابل بوجود میآیند. گر چه از نظر اندازه هر باز پورینی میتواند در مقابل یک باز پیریمیدین قرار بگیرد. ولی به دلیل وجود گروههای شیمیایی روی بازهای G و C و T و A پیوندهای هیدروژنی مناسب فقط بین C - G و T - A برقرار میشود و ایجاد پیوند بین T - G و C- A ممکن نیست.
واکنشهای توتومریزاسیون
اتم هیدروژن در بازهای آلی میتواند روی اتمهای نیتروژن و یا اکسیژن حلقه جابجا شود. این تغییر موقعیت هیدروژن روی حلقه باز را توتومریزاسیون میگویند. توتومریزاسیون در بازهای آدنین سیتوزین باعث تبدیل فرم آمینی به فرم ایمنی و در مورد بازهای تیمین و گوانین باعث تبدیل فرم کتونی به فرم انولی میشود.
در شرایط فیزیولوژیکی ثابت تعادل واکنش توتومریزاسیون بیشتر به سمت اشکال آمینی و کتونی میباشد. این حالت پایدار پروتونی ، الگوی تشکل پیوندهای هیدروژنی بین بازها را تعیین مینماید، بطوری که بازهای T و A با تشکیل دو پیوند هیدروژنی و بازهای G و C با سه پیوند هیدروژنی با هم جفت میشوند. C و A و همچنین T و G نمیتوانند با هم جفت شوند.
زیرا در این بازها اتمهای هیدروژن هر دو در یک موقعیت قرار دارند و امکان ایجاد پیوند هیدروژنی وجود ندارد. به دلیل اینکه در رشتههای DNA همواره باز A مقابل T و باز G مقابل C قرار دارد، این دو رشته را مکمل مینامند. بنابراین توالی موجود در یک رشته DNA ، توالی رشته مقابل را تعیین میکند. مکمل بودن دو رشته DNA ، اساس عمل همانند سازی DNA است.
مقدمه ای بر تقسیم میوز
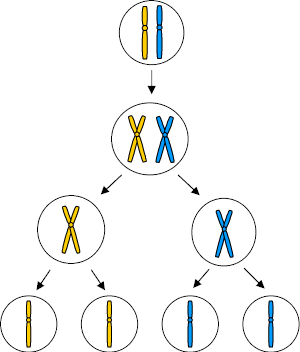
بخش اول میوز همانند میتوز خود شامل چهار مرحله است.
مرحله پروفاز در میوز اول روند پیچیدهای است که بسیار کندتر از میتوز صورت میگیرد و شامل پنج مرحله است:
-
زیرمرحله لپتوتن:
آغاز پروفاز با افزایش حجم هستهای مشخص میشود. کروموزومها به صورت تخمهای دراز ، نازک و تاب خورده به شکل دانههای تسبیح به نام کرومومر ظاهر میشوند. این ریز مرحله را لپتوتن گویند. کروموزومها منفرد به نظر میرسند، در حالی که بیشتر DNAی یاخته قبلا دو برابر شده و کروموزومها دارای دو کروماتید هستند. بر اساس گفته «براون» ، سنتز DNA تا مرحله لپتوتن ادامه دارد و زمان چرخه یاختهای را تشکیل میدهد.
چرخه یاختهای را تشکیل میدهد. -
زیرمرحله زیگوتن:
در این مرحله کروموزومهای همساخت به ترتیب ویژهای جفت میشوند. نیرویی که دو جفت کروموزوم را به سوی یکدیگر میکشد، هنوز مشخص نشده است. این روند را سیناپس میگویند و جفت کروموزومهای همساخت را بیوالانت (تتراد) میگویند. -
زیرمرحله پاکیتن:
در این مرحله هستک از نظر اندازه رشد میکند و کروموزومها کوتاهتر و ضخیخمتر میشوند. حال هر کدام یک تتراد هستند که از دو کروموزوم همساخت یا 4 کروماتید تشکیل شدهاند. هر کروماتید از یک تتراد ، به دور کروماتید خواهر خود میپیچد و کوتاهتر و ضخیمتر میشود. هر کروموزوم همساخت سانترومر مستقل دارد. بنابراین هر کروماتید سانترومر خاص خود را دارا است.
مهمترین رویداد در زیرمرحله پاکیتن ، تشکیل کیاسما به هنگامی است که دو کروماتید خواهر از هر کروموزوم همساخت ، قطعاتی را بین خود مبادله میکنند. تبادل قطعات بین دو کروماتید از دو کروموزوم همساخت را کراسینگ اور (تقاطع کروموزومی) گویند. زیرمرحله پاکیتن طولانی است. در پایان این زیرمرحله ، نیرویی سبب جدا شدن کروماتیدها از یکدیگر میشود. -
زیرمرحله دیپلوتن:
در این مرحله کروموزومها ، جدا شدن از یکدیگر را آغاز میکنند، اما چون در بعضی نقاط تبادل صورت گرفته است، لذا در این نقاط متصل به یکدیگر باقی میمانند. این ریز مرحله حقیقتا کیاسما نام دارد و از نظر ژنتیکی دارای اهمیت فراوانی است، زیرا تبادل بین کروماتیدهای ناخواهری در این زیرمرحله صورت میگیرد. کراسینگ اور به تبادل ژنها میانجامد و سبب تشکیل کروماتیدهای نوترکیب میشود. در ژنتیک مولکولی ، کراسینگ اور به عنوان وسیله تجربی برای نقشه برداری کروموزومی بکار میرود. -
زیرمرحله دیاکینز:
در این مرحله ، کروموزومها کوتاهتر و ضخیمتر شده و کیاسما ناپدید میشود. کروموزومهای همساخت از دو سو به سمت محیط هسته کشیده میشوند، اما جدا شدن کامل کروماتیدها صورت نمیگیرد. کروموزومهای همساخت فقط در انتها متصل به یکدیگر باقی میمانند و ساختار حلقه مانند عریضی را تشکیل میدهند. به علاوه هستک و غشای هسته ناپدید میشود و دوک بطور کامل تشکیل میگردد. کرومزومهای تتراد در صفحه متافاز قرار میگیرند.
متافاز اول
این مرحله پس از دیاکینز آغاز میشود و همانند متافاز میتوز است. کروموزومهای همساخت در صفحه استوایی باقی میمانند و از طریق سانترومرها به رشتههای دوک متصل میشوند.
آنافاز اول
در آنافاز اول ، کروماتیدهای خواهر از هر کروموزوم همساخت که به وسیله سانترومر به یکدیگر متصلاند، به قطبهای مربوط به خود میروند. کیاسما کاملا متلاشی میشود و کروماتیدهای ناخواهری از هم جدا میگردند. این کروماتیدها ، با کروموزومهای پدری و مادری خود تفاوت دارند. در مقایسه با آنافاز میتوز که در آن هر کروموزوم یک کروماتید دارد، هر کروموزوم در مرحله آنافاز میوز ، از دو کروماتید تشکیل شده است که احتمالا یکی از کروماتیدها ، نوترکیب است.
تلوفاز اول
در این مرحله کوتاه ، پیچش کروماتیدها باز شده و کروماتیدها دراز میشوند و تا مدتی در حالت فشردگی باقی میمانند. غشای هسته در اطراف هر گروه کروماتید تشکیل میگردد و دو هسته مجزا بوجود میآیند. در بعضی موجودات پس از تشکیل غشاها در هسته ، هر هسته دختر قبل از اینکه دومین تقسیم میوز آغاز شود، مدتی در مرحله اینترفاز باقی میماند. باید توجه داشت که بین دو تقسیم میوز (ساختمان DNA|DNA)) ساخته نمیشود. 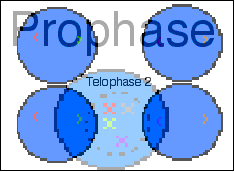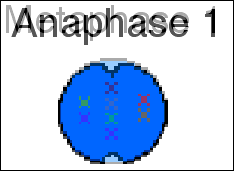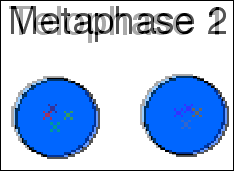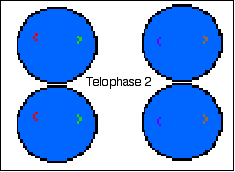
مرحله دوم میوز
این مرحله تقسیم همانند میتوز است، اما با این تفاوت که کروموزومها از دو کروماتید تشکیل شدهاند. در این نوع تقسیم هر دو هسته خواهر از مراحل پروفاز ، متافاز ، آنافاز و تلوفاز دوم میگذرند. در این مرحله مضاعف شدن DNA صورت نمیگیرد.
پروفاز دوم
پروفاز این مرحله بسیار کوتاه است. دوک تشکیل میشود و کروموزومهای دو کروماتیدی و مضاعف روی آن قرار میگیرند.
متافاز دوم
در متافاز دوم ، کروموزومها به قسمت وسط دوک میروند و در آنجا مستقر میشوند. نکته جالب توجه این است در متافاز میوز اول سانترومرهای کروموزومهای همساخت از یکدیگر جدا میشوند، در حالی که در میوز دوم سانترومرهای کروماتیدهای خواهری از یکدیگر فاصله میگیرند.
آنافاز دوم
در آنافاز دوم میوز کروماتیدهای هر کروموزوم از هم جدا میشوند و به دو قطب سلول میروند.
تلوفاز دوم
در تلوفاز دوم میوز ، تقسیم میوزی کامل میشود و چهار سلول بوجود میآید. در بسیاری از جانداران ماده ، سیتوپلاسم سلولها در میوز بطور نامساوی تقسیم میشود و فقط یک سلول به جای چهار سلول حاصل میآید که سیتوپلاسم فراوان دارد و مبدل به تخمک میشود. سه سلول کوچک باقیمانده معمولا میمیرند. در بعضی از جانداران نر چهار سلول حاصل مبدل به اسپرم میشوند.
بکرزایی
در هر جاندار ، اعم از اینکه تک جنسی باشد یا هرمافرودیت ، بسیار اتفاق میافتد که نتوانند جفت مناسبی پیدا کنند. بیشتر چنین گامتهای ناموفقی پس از اندک زمانی متلاشی میشوند. اما در موارد استثنایی ، گامتهای منفرد میتوانند رشد کنند و افراد بالغ طبیعی بوجود آورند. این پدیده را بکرزایی (Parthenogenesis) گویند. از آنجا که هیچ جاندار نری در این نوع تولید مثل شرکت ندارد تا ماده ژنتیک خود را به اشتراک بگذارد، فرزندان حاصل کاملا شبیه مادر هستند و در واقع یک کلون هستند. در طبیعت کلونها در نتیجه تولید مثل غیر جنسی پدید میآیند. بکرزایی ، شکل ویژهای از کلون کردن است که به انواع تولید مثل تعلق دارد. 
بکرزایی در چه جاندارانی رخ میدهد؟
در بسیاری از آغازیان ، هر نوع گامت میتواند به شیوه بکرزایی رشد کند. در جانوران ائوگام ، فقط تخمکها گاهی میتوانند به این شیوه رشد کنند. از اینرو ، بکرزایی طبیعی مثلا در روتیفرها ، زنبورها و دیگر حشرات اجتماعی صورت میگیرد. و در پرندگانی از قبیل بوقلمون و مرغ خانگی نیز بندرت رخ میدهد. آپومیکسی نوعی بکرزایی است که در گیاهان رخ میدهد. در این پدیده ، جنین از طریق لقاح حاصل نمیشود. بلکه از یاختههای داخل کیسه جنینی و یا یاختههای خورش اطراف کیسه جنینی بوجود میآید. در بکرزایی گامتی که قادر به بکرزایی است، از نظر کنش تفاوتی با هاگ ندارد یعنی هر دوی آنها سلولهای زایندهای هستند که میتوانند مستقیما به فرد بالغ تبدیل شوند.
بکرزایی طبیعی
حشرات اجتماعی از جمله زنبورها ، جانورانی هستند که در آنها پدیده بکرزایی اتفاق میافتد. به این ترتیب که ، هنگامی جمعیت یک کندو زیاده از حد شود. ملکه و چند زنبور نر و چندین هزار کارگر از کلونی جدا میشوند. ملکه و زنبور نر با هم جفت تشکیل میدهند. اسپرمهای دریافت شده توسط ملکه در کیسهای واقع در درون شکم ذخیره میشوند. اسپرمهای دریافت شده از همین یکبار جفتگیری تا آخر دوره تخمگذاری ملکه باقی میمانند.
در میان تخمهایی که یکی یکی در جایگاههای شانه کندو زنبور نهاده میشوند، حتی در بدن ملکه جوان هم بعضی از آنها بارور نمیشوند. در ملکه پیری که ذخیره اسپرم خود را به پایان رسانده ، هیچکدام از تخمها بارور نمیشوند. تخمهای بارور نشده ، زنبورهای نر را میسازند. چنین رشد محروم از پدری را که معمولا در میان حشرات اجتماعی فراوان است، بکرزایی طبیعی گویند.
بکرزایی مصنوعی
در بعضی از گونههای جانداران ، با استفاده از وسایل آزمایشگاهی ، بکرزایی مصنوعی ایجاد میکنند. مثلا میتوان با فرو کردن سوزن بر سطح تخمک بارور نشده قورباغه (همراه با تدابیر دیگر) ، آن را وادار به رشد کرد. نیش سوزن بر سطح تخمک ، اثری چون نفوذ اسپرم دارد و رشد آن را سبب میشود. سرد کردن تخمک بارور نشده یا تحریک کردن آن با اسیدهای قوی از دیگر راههای بکرزایی مصنوعی به شمار میروند.
بکرزایی در روتیفرها
رده روتیفرها جز شاخه آشلمنتها یا کرم سانان رده بندی میشوند و شامل 1500 گونه هستند. روتیفرهای ماده در طول بهار و تابستان تخمکهایی تولید میکنند که بدون لقاح ، روتیفرهای ماده دیگری را پدید میآورند. این مادهها نیز بدون لقاح تولید مثل میکنند و به این روش ، نسلهای ماده متوالی حاصل میآید. در پاییز ، مادهها چند تخمی کوچکتر از بقیه میگذارند که از آنها نرهای کوچکی حاصل میآیند.
نرها میتوانند اسپرم تولید کنند که با تخمک لقاح یافته و تخم حاصل میشود که دارای پوستهای ضخیم است که میتواند شرایط نامساعد را تحمل کند. در شرایط مساعد مثلا در بهار سال بعد ، از این تخمها ، کرمهای ماده حاصل میشود. در بعضی از اقسام روتیفرها ، اصولا جنس نر دیده نمیشود و افراد همگی حاصل از رشد تخمکهای لقاح نیافته هستند که رشد بدون لقاح یا بکرزایی است. 
بکرزایی در گیاهان
بذرهای غیر جنسی یا نامیزه (Ampomict) در اثر پدیده آپومیکسی حاصل میشوند. پدیده آپومیکسی نوعی بکرزایی بشمار میرود و جنینها در اثر بی نر زایی (Agamosprmy) حاصل میشوند. در پدیده آپومیکسی ، تولید مثل غیر جنسی جانشین تولید مثل جنسی در اندامهای اختصاص یافته به تولید مثل جنسی میگردد.
آپومیکسی مداوم
در این حالت ، کیسه جنینی از سلول مادری تخم حاصل میشود و دیپلوئید میباشد. اما سلول مادری تخم تقسیم میوز ناقص انجام میدهد و در نتیجه سلول تخم همانند گیاه مادری دیپلوئید بوده و جنین غیرجنسی نیز که دیپلوئید میباشد بطور مستقیم از هسته تخم و بدون عمل لقاح حاصل میشود. این حالت در گیاهانی مانند پیاز ، گل قاصدک ، سیب و تمشک دیده میشود.
بکرزایی هاپلوئید
این پدیده را آپومیکسی غیر مداوم نیز مینامند. در این حالت جنین غیر جنسی از سلول تخم زا که هاپلوئید است، بوجود میآید و نهال تولید شده نیز هاپلوئید خواهد بود. این پدیده بندرت افتاق میافتد. برای مثال در 25 هزار نهال هلو احتمال دارد که 4-3 نهال هاپلوئید مشاهده شود.
آپوگامی
در این حالت جنینهای غیر جنسی از دو سلول قرینه در کیسه جنینی مثلا در سوسن ، و یا جنین هاپلوئید از سلولهای متقاطر در کیسه جنینی مثلا در سیاهدانه حاصل میشوند.
اهمیت پدیده بکرزایی در گیاهان
از آنجایی که اکثر گیاهان تولید شده در پدیده آپومیکسی زا سلولهای پایه مادری میباشند، در نتیجه گیاهان بوجود آمده شبیه پایه مادری میباشند. در این نوع گیاهان ، تفرقه صفات کمتر مشاهده میشود. بذور غیر جنسی نیر شبیه بذرهای جنسی مراحل مختلف نونهالی ، انتقال و بلوغ را سپری میکنند. بذرهای حاصل از بکرزایی سریعتر رشد میکنند. از آنجایی که این بذرها معمولا فاقد ویروس هستند، در نتیجه نهالهای تولید شده ، عاری از ویروس خواهند بود.
